

Abstract
Glutamate is the most abundant excitatory neurotransmitter in the brain, and it plays an essential and important role in neural functions. Hypofunction of the glutamatergic pathway and the changes in the glutamate-glutamine cycle function are important neuropathological mechanisms of severe mental disorders including schizophrenia and depression. Current studies have shown that glutamate can induce neural biophotonic activity and transmission, which may involve the mechanism of photon quantum brain; however, it is unclear whether such a mechanism follows the principle of quantum mechanics. Here we show that the action of glutamate on its receptors leads to a decrease in its quantum energy levels, and glutamate then partially or completely loses its function to further induce the biophotonic activity in mouse brain slices. The reduced quantum energy levels of glutamate can be restored by direct-current electrical discharges and the use of energy transfer of chloroplast photosynthesis; hence, the quantum energy recovered glutamate can again induce significant biophotonic activity. Furthermore, the changes in quantum energy levels of glutamate are related to the exchange and transfer of electron energy on its active hydrogen atom. These findings suggest that the glutamate-induced neural biophotonic signals may be involved in the transfer of the quantum energy levels of glutamate, which implies a quantum mechanism of neurotransmitter action. The process of glutamate recycling that is related to the synergism of neurons and glial cells and certain key enzymes may be necessary for the recovery of quantum energy levels of glutamate after completion of the neural signal transmission. These findings may also provide a new idea to develop “quantum drugs”.
Introduction
Analysis of the binding characteristics of biological signaling molecules to their targets or receptors provides an important means to understand their physiological roles and to develop target drugs. However, a large number of studies have found that the effects of signaling molecules on their targets are often unpredictable, for example, the signal molecules or drugs with identical or similar structures such as chiral drugs may produce different effects, while the molecules with completely different structures may induce identical or similar effects (1). For olfactory molecules, human beings can directly experience their differences, and quite different molecules can have similar odors, whereas similar molecules can have dissimilar odors (2).
The theoretical studies speculate that the reasons for these phenomena may be related to the mechanism of quantum biological action (3-5), however, the theoretical framework based on traditional quantum mechanics, such as quantum coherence, entanglement and superposition, could not explain the existence of phenomena very well (6, 7). Recent studies have shown that biophoton activity may play an important role in quantum biological mechanisms, and it was reported that glutamate, the most abundant excitatory neurotransmitter in the brain, could induce biophotonic activities and transmission in neural circuits, which may involve the mechanism of photon quantum brain (8-12). The characteristic differences such as spectral redshift in glutamate-induced biophotonic activity in different animals may give some evolutionary advantages such as human intelligence (13), but may also allow human beings to be more sensitive to the development of neurological and psychiatric diseases due to the changes in neurotransmitter functions. For example, hypofunction of the glutamatergic pathway and the changes in the glutamate-glutamine cycle function are important neuropathological mechanisms of severe mental disorders including schizophrenia and depression (14-21). Therefore, we hypothesize that the enhancement or decrease of glutamate function may be related to the changes in quantum energy levels of glutamate, which were experimentally investigated in this study.
Results
NMDA receptor mediates glutamate-induced biophotonic activity
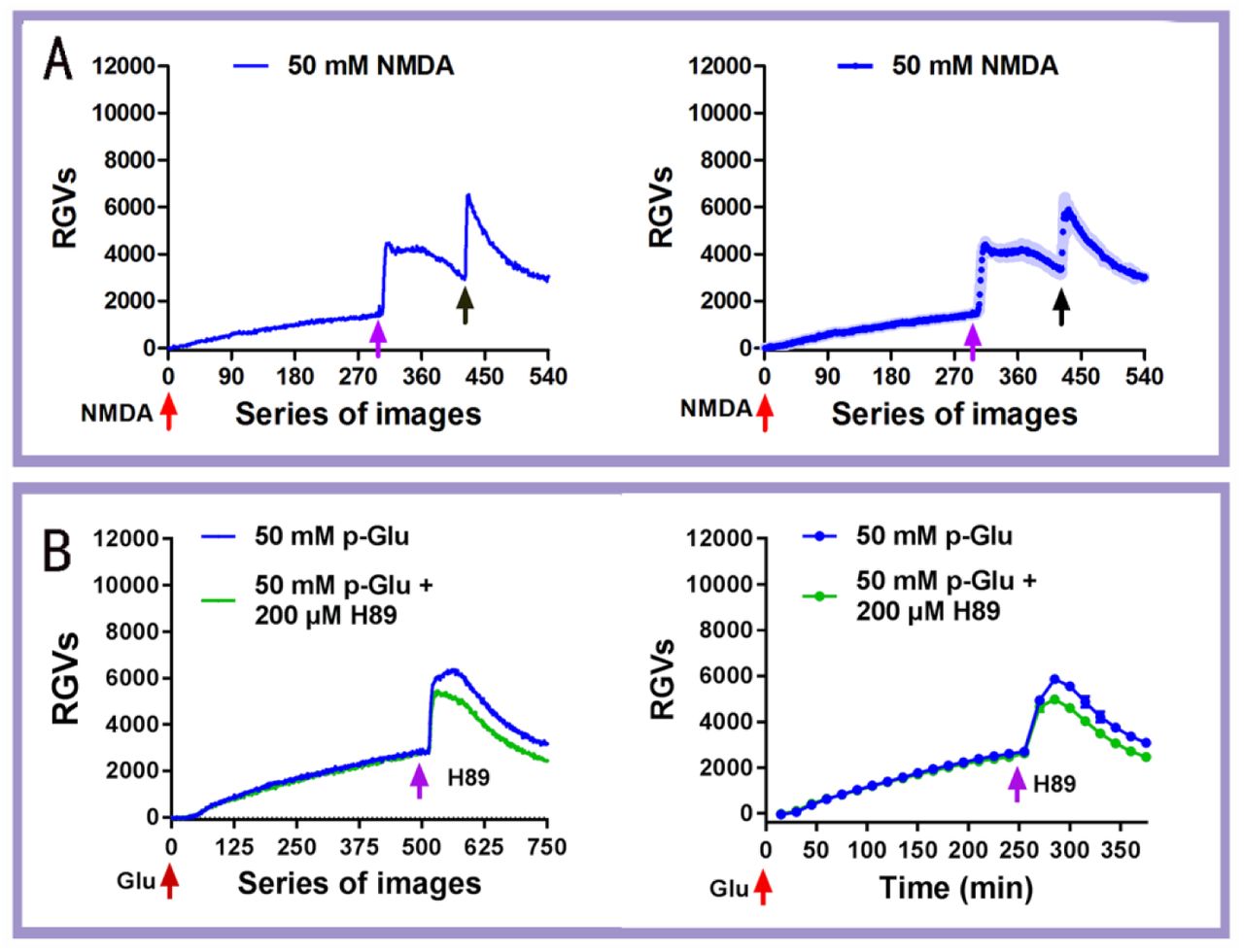
Our previous study demonstrated that 50 mM purchased glutamate (p-Glu) can lead to biophotonic activity and transmission (neural biophotonic signals) in neural circuits, presenting four characteristic stages (initiation, maintenance, washing and reapplication) (8). In this study, we further found that N-Methyl-D-aspartate (NMDA), a specific agonist at one of the ionotropic glutamate receptors, could also induce similar patterns of biophotonic activity in mouse brain slices (Fig. 1A), suggesting that the NMDA receptor mediates glutamate-induced biophotonic activity. In addition, the washing effect, which provides a further increase in biophotonic activity, was significantly inhibited by the application of a presynaptic vesicle release inhibitor (H89) at the beginning of washing (Fig. 1B and Table S1), suggesting that the washing effect is due to the enhanced presynaptic glutamate release. A possible mechanism is that the extracellular application of glutamate can reduce the release of presynaptic vesicles through an inhibitory effect on the presynaptic membrane (22), which results in increased glutamate storage in the presynaptic vesicles and, therefore, a rapid and massive glutamate release from the presynaptic membrane occurs during the early stage of washing.

(A) Representative dynamic changes in biophotonic activity demonstrated by the relative grey values (RGVs) in the left panel after the application of 50 mM NMDA, showing four characteristic stages (initiation, maintenance, washing and reapplication; 30 s imaging time for each original image). The population average dynamic changes in biophotonic activity demonstrated by RGVs are in the right panel (n=5). Shaded regions indicate s.e.m. values. The red, pink and black arrows indicate the time points for the application (0 min), washing (150 min) and reapplication (210 min) of NMDA, respectively.
(B) Representative dynamic changes in biophotonic activity demonstrated by the RGVs in the left panel, showing three characteristic stages (initiation, maintenance and washing; 30 s imaging time for each original image). The right panel shows the sum of the time course of the average change in RGVs from 15 continuously processed original grey images. The red and pink arrows indicate the time points for the application of glutamate (0 min) and 200 μM H89 (240 min) just before washing. The intensities of biophotonic activity tended to decrease significantly after the application of H89 compared to those in controls (n=6 for 50 mM p-Glu and 50 mM p-Glu+200 μM H89).
Data are shown as the mean±s. e. m. n=the number of slices from the same number of mice. Significant differences (** p<0.01) are noted from 271-285 min (see Table S1).
Decrease in the quantum energy levels of glutamate after its action
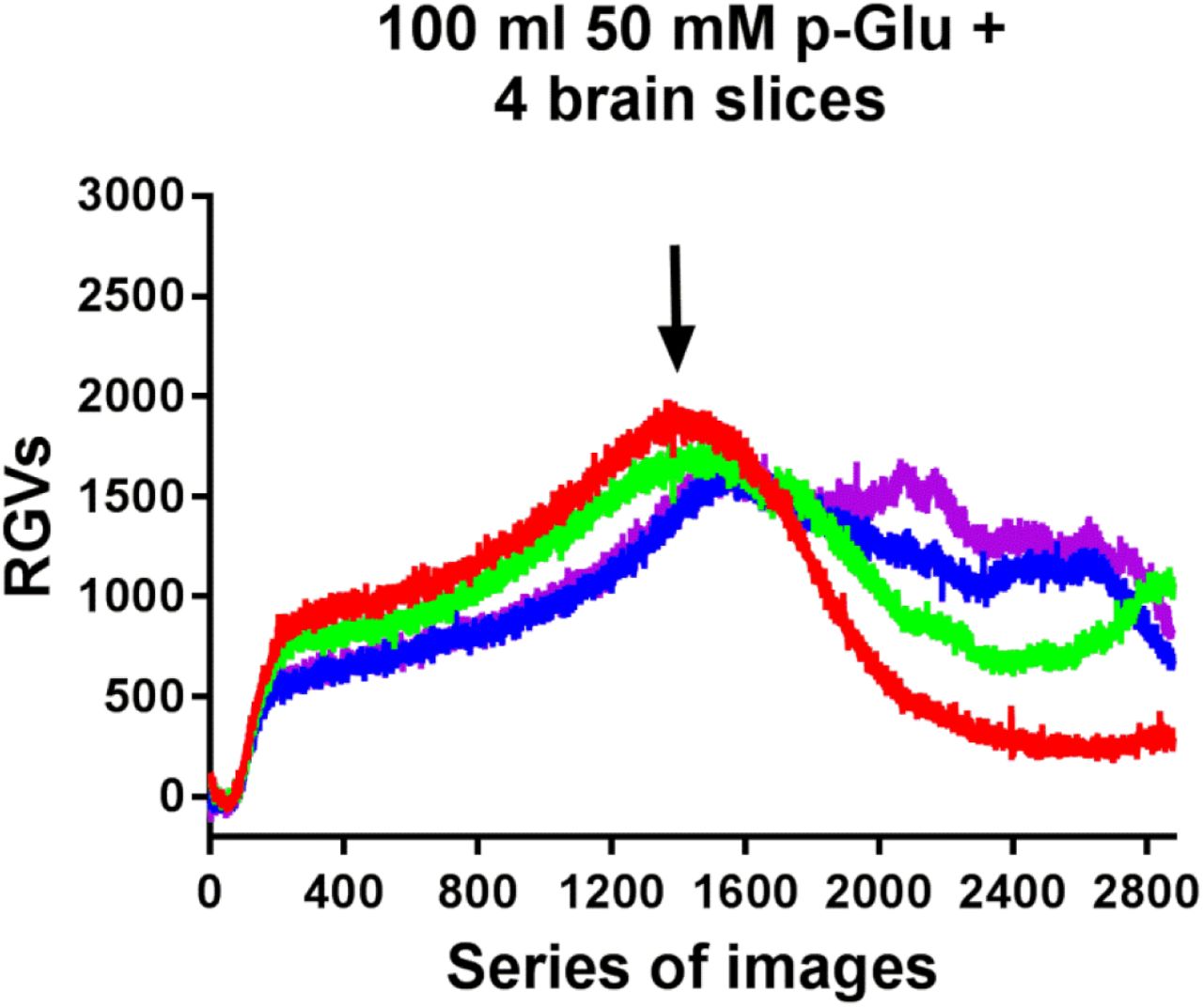
Then, we observed long-term (24 h) effects of 50 mM glutamate-induced biophotonic activity. We designed a consumption experiment to evaluate the changing patterns of glutamate-induced biophotonic activity. Mouse brain coronal slices (450 µm in thickness) were cut from the anterior part of the olfactory cortex to the caudal part of cerebellar cortex and all slices were collected. We found that the intensities of glutamate-induced biophotonic activities from a slice for imaging, which was perfused with 100 ml ACSF containing 50 mM glutamate together with four whole mouse brain slices in a bottle, dramatically decreased after perfusion and imaging for approximately 12 h (12.02±0.65 h, Fig. S1). This result allowed us to speculate that the cause of the obvious weakening of glutamate action may be related to the decrease in the quantum energy levels of glutamate after its action on the postsynaptic receptors (Fig. 2A) because a cyclic perfusion with a fixed volume of ACSF containing 50 mM glutamate was able to partially and almost completely exhaust the action of glutamate by providing appropriate brain tissues and enough perfusion time.

Dynamic changes in the biophotonic activity in an imaging slice demonstrated by RGVs are shown after the cyclic perfusion with 100 ml of ACSF containing 50 mM p-Glu and four whole mouse brain slices, which were placed into a perfusion bottle. The intensities of biophotonic activity in four imaging slices (red, green, blue and pink, 30 s imaging time for each image) tended to decrease significantly after approximately 12 h of perfusion (12.02±0.65, n=4). The total imaging time was 24 h for each case.

(A)Schematic drawing of the decrease in quantum energy levels from the level at the normal competent (Nc) to the levels of partial and complete loss of quantum energy after the glutamate action on its receptors. Ic: incompetent; p-Ic: partial incompetent.
(B) Schematic drawing of cyclic dual perfusion system, named perfusion system A and B. The perfusion chamber under the same imaging field is separated into two smaller perfusion parts (subchambers) (see the figure, the right part of the panel), allowing support of independent cyclic perfusion for the left and right hemisphere slices together with two perfusion solution storage glass bottles (X and Y) and input and output micropumps.
(C, D) Representative dynamic changes in biophotonic activity demonstrated by the RGVs are shown in the upper left panel. The sum of the time course of the average change in RGVs from 30 continuously processed original grey images is shown in the upper right panel. One of the two hemisphere slices (lower slice in the lower left regular photograph) was perfused independently with 50 ml of ACSF containing 50 mM p-Glu after treatment with 2 whole brain slices (C) or 4 whole brain slices (D) for 24 h. Another one (upper slice in the lower left regular photograph) was also perfused independently with 50 ml of ACSF containing 50 mM p-Glu as a control. Representative biophoton grey images at the selected time periods indicated in the lower panel and digit: 1-4 mark the 90, 270, 360 and 450 series of images, respectively (30 s imaging time for each image). The red, pink and black arrows indicate the time points for application of perfusion solution (0 min), washing (150 min) and reapplication (210 min), respectively. The intensities of biophotonic activity were decreased significantly after treatment with 2 whole brain slices for 24 h (green lines) compared to those of the controls (blue lines, c, n=6), and were almost completely eliminated with 4 whole brain slices (D, n=6). p-Glu: purchased glutamate.
Data are shown as the mean±s. e. m. n=the number of slices from the same number of mice. Significant differences (* p<0.05 or ** p<0.01) are noted from 91-105 min for C and 1-15 min for D (see also Table S1).
To confirm our results, we designed a cyclic dual perfusion system to conduct further experiments. Two hemisphere slices (left and right, 450 µm in thickness) prepared from a coronal whole brain slice at the anterior level of the hippocampus were placed separately into two perfusion subchambers that were imaged simultaneously (Fig. 2B). We found that the patterns and intensities of 50 mM glutamate-induced biophotonic activity were exactly the same for both slices (Fig. S2), which was consistent with data obtained from a whole brain coronal slice containing left and right hemispheres (8). Then, we tested the effects by only using ACSF containing 50 mM p-Glu after incubation with different quantities of brain slices. All slices prepared from two or four mouse brains were incubated in 100 ml ACSF containing 50 mM p-Glu for 24 h at ~ 28℃. Then, 50 ml incubated solution was collected to perfuse a new prepared brain slice for biophotonic imaging. We found that the intensity of biophotonic activity during the four stages induced by the solution after incubation with two whole brain slices was significantly weaker than that in control solution (Fig. 2C and Table S1), while the effects were almost eliminated after incubation with four whole brain slices, both of which resulted in hypofunctional (partial incompetent, p-Ic) and nonfunctional glutamate or incompetent glutamate (Ic-Glu) (Fig. 2D and Table S1). The observed results were not due to the change in the molecular structure of glutamate after the action of its receptors or the effects of other active molecules released from the brain slices during incubation. Even if the low concentration of classical neurotransmitters such as acetylcholine, dopamine, 5-hydroxytryptamine (5-HT) and norepinephrine were present in the ACSF after incubation for 24 h, their presence cannot explain the weakening and disappearance of glutamate action because our previous study demonstrated that even high concentrations of 5-HT (> 20 µM) only produced partial inhibitory effects on 50 mM glutamate-induced biophotonic activity, while acetylcholine, dopamine and norepinephrine played reinforcing rather than inhibitory roles (12). Therefore, in essence, these results proved our speculation mentioned above.

Representative dynamic changes in the biophotonic activity demonstrated by RGVs are shown in the upper left panel after the cyclic perfusion of left and right hemisphere slices with 50 ml ACSF containing 50 mM glutamate. The sum of the time course of the average change in RGVs from 30 continuously processed original grey images is shown in the upper right panel (n=10). Red and blue lines indicate the dynamic biophotonic activity in the up and lower slices, respectively. The patterns and intensities of 50 mM glutamate-induced biophotonic activity were the same for both slices, which was consistent with that obtained from a whole brain slice containing left and right hemispheres. Digit: 1-4 mark the 90, 270, 360, and 450 series of images, respectively (30 s imaging time for each image) in the upper left panel, which corresponds to the four images in down panel. The red, pink and black arrows indicate the time points for application of 50 mM glutamate (0 min), washing (150 min) and reapplication (210 min), respectively. A representative regular image of two brain slices (up and down) is shown on the left side of the lower panel.
Data are shown as the mean±s. e. m. n=the number of slices from the same number of mice.
Electrical discharge restores the quantum energy levels and the effects of incompetent glutamate
To test whether there are ways to restore the decreased quantum energy levels of glutamate and reverse its action and function to induce biophotonic activity, we developed two techniques (Fig. 3A). One is called the electrical stimulation assisted quantum energy enhancement technique (ESA-QEET, Fig. 3B, see also SI for details). One hundred millilitres ACSF containing 50 mM p-Glu was first incubated with four whole brain slices for 24 h at ~ 28°C and then divided into two parts (50 ml each). One portion of the solution was treated for 3 h with 12 V direct-current electrical discharges (DC-ED) at a high frequency (~10 k Hz), and another was the control. We found that such treatments significantly restored the biophotonic activity of Ic-Glu compared to the controls without DC-ED treatment and reached to 59.00-118.25% of the effects of ACSF containing 50 mM p-Glu at the different imaging periods (Fig. 3C and Table S1). The recovery of the quantum energy level of glutamate was not due to the influence of DC-ED on the other components of the ACSF because the ACSF without p-Glu incubated with four whole brain slices for 24 h did not have any effects on the biophotonic activity after treatment with 12 V DC-ED (Fig. 3D and Table S1). These findings also imply that the increase in quantum energy levels of glutamate may be involved in the transfer of electron energy on its molecule, and a possible explanation is that the active hydrogen (proton) of glutamate is responsible for the exchange and transfer of quantum energy, which was supported by our findings that, similar to 50 mM p-Glu, 50 mM acetic acid (0.3%) could also induce similar patterns of biophotonic activity in mouse brain slices (Fig. S3 and Table S1), whereas 50 mM sodium glutamate (gourmet powder), on which the sodium atom is substituted for the active hydrogen atom, could not have any effect on the induction of biophotonic activity in mouse brain slices (Fig. S4 and Table S1).

Representative dynamic changes in the biophotonic activity demonstrated by RGVs are shown in left panel (30 s imaging time for each image). The sum of the time course of the average change in RGVs from 15 continuously processed original grey images is shown in right panel. Digit: 1-4 mark the 90, 270, 360, and 450 series of images, respectively, in the upper left panel, which corresponds to the four images in down panel. The red, pink and black arrows indicate the time points for application of 50 mM glutamate (upper image) and 50 mM acetic acid (0.3%, lower image) (0 min), washing (150 min) and reapplication (210 min), respectively. A representative regular image of two hemispherical slices (up and down) is shown on the left side of the lower panel. Acetic acid (50 mM) (red line, down image) also induced similar patterns of biophotonic activity compared to that of 50 mM p-Glu (blue line, upper image) (n=5).
Data are shown as the mean±s. e. m. n=the number of slices from the same number of mice.

Representative dynamic changes in the biophotonic activity demonstrated by RGVs are shown in the left panel (30 s imaging time for each image). The sum of the time course of the average change in RGVs from 30 continuously processed original grey images is shown in the right panel. Digit: 1-4 mark the 90, 270, 360, and 450 series of images, respectively, in upper left panel, which corresponds to the four images in down panel. The red, pink and black arrows indicate the time points for application of 50 mM glutamate (upper image) or 50 mM sodium glutamate (down image) (0 min), washing (150 min) and reapplication (210 min), respectively. A representative regular image of two hemispherical slices (up and down) is shown on the left side of the lower panel. Sodium glutamate (50 mM) (red line, down image) did not induce biophotonic activity, while 50 mM p-Glu (blue line, upper image) had typical effects (n=5).
Data are shown as the mean±s. e. m. n=the number of slices from the same number of mice. Significant differences (* p<0.05 or ** p<0.01) are noted from 1-15 min (see also table S1).

(A) Schematic drawing of the potential recovery of the quantum energy levels (Rc) of glutamate after treatment with the electrical stimulation assisted quantum energy enhancement technique (ESA-QEET, lower red arrow) or chloroplast-assisted quantum energy enhancement technique (CA-QEET). The levels at super competent (Sc) can also be obtained by ESA-QEET from the normal level (upper red arrow) or by CA-QEET from the incompetent level (blue arrow).
(B) Schematic drawing of the ESA-QEET. A power supply device with 12 V direct-current electrical discharges (DC-ED) at a high frequency (~10 k Hz) was used to execute the electrical stimulations using the two same carbon rods as the cathode and anode.
(C, D) Representative dynamic changes in the biophotonic activity demonstrated by the RGVs are shown in the upper left panel. The sum of the time course of the average change in RGVs from 30 continuously processed original grey images is shown in the upper right panel. One of two hemisphere slices (upper slice in the lower left regular photograph) was perfused independently with the recompetent glutamate treated by 12 V DC-ED for 3 h after its incompetent treatment with 4 whole brain slices for 24 h (C) or ACSF treated by 12 V DC-ED for 3 h after incubation with 4 whole mouse brain slices for 24 h (D). Another hemisphere slice (lower slice in the lower left regular photograph) was perfused independently with the incompetent glutamate without DC-ED treatment (C) or ACSF without treatment by 12 V DC-ED after incubation with 4 whole mouse brain slices for 24 h (D) and considered the controls. Representative biophoton grey images at the selected time periods indicated in the upper left panel and digit: 1-4 mark the 90, 270, 360 and 450 series of images, respectively (30 s imaging time for each image). The red, pink and black arrows indicate the time points for application of the perfusion solution (0 min), washing (150 min) and reapplication (210 min), respectively. Approximately 59.00-118.25% of the effects of p-Glu (red line) were reached at the different time imaging periods after treatment for 3 h with 12 V DC-ED (n=8) compared to the reference induced by 50 mM p-Glu during the different imaging periods (n=37) (C). No significant increase in biophotonic activity in four stages after treatment with DC-ED (n=5) was observed compared to that without treatment (D). p-Glu: purchased glutamate.
Data are shown as the mean±s. e. m. n= the number of slices from the same number of mice. Significant differences (* p<0.05 or ** p<0.01) are noted from 1-15 min for c (see also Table S1).
Chloroplast photosynthesis restores the quantum energy levels and the effects of incompetent glutamate
Another technique is called the chloroplast-assisted quantum energy enhancement technique (CA-QEET, Fig. 3A, see also SI for details). Chloroplasts can achieve efficient light energy transfer through photosynthesis under light illumination and, therefore, the electron and proton transport between molecules can be realized, in which quantum mechanics is involved (23-26). We explored whether chloroplasts were able to allow the hypofunctional or nonfunctional glutamate to restore its quantum energy levels. Chloroplasts were prepared from a vegetable (spinach) and a comparative analysis showed that the isolated chloroplasts could maintain a certain degree of light transfer activity in the ACSF and the ACSF containing 50 mM p-Glu compared to those under an appropriate condition reported previously (27) (Fig. S5). We added ~3 g freshly prepared chloroplast precipitate into 50 ml ACSF containing Ic-Glu in two steps (~1.5 g each) and incubated the mixed preparations for 48 h under a combination of day and night illumination (see SI for details). We found that such treatments allowed Ic-Glu to reach 160.25-370.70% of the effects of the 50 mM p-Glu at the different imaging periods (Fig. 4A and Table S2), showing the super recompetent effects (Fig. 3A). The over-recovery of biophotonic activity was partially because of the acidification effect of light energy transfer of the chloroplasts on ACSF (28-30), which resulted in low pH (from ~7.4 to ~4.0) and induced some degree of biophotonic activity both from the ACSF treated with and without four whole brain slices (Fig. 4B and Table S2); however, adjusting the acidified ACSF to the original level of pH 7.4 significantly reduced the effects and only induced slight biophotonic activity (Fig. 4C and Table S2), reinforcing the role of the active hydrogen for the glutamate-induced biophotonic activity mentioned above.

(A-P) Three reaction solutions (50 ml each) including ACSF, ACSF containing 50 mM p-Glu and reaction mixture [50 mM Tris-HCl buffer (pH 7.4), 0.4 mM sucrose, 10 mM NaC1] were placed into the left, middle and right bottles, respectively.
(A) Freshly prepared chloroplast precipitate (1.5g) was added into the three reaction solutions (dark green).
(B) One milliliter of 50 mM DCIP was added at 13:41 (highlights in red, dark blue).
(C-I) The colour change from dark blue to purple-red-blue was observed for ACSF containing 50 mM p-Glu after incubation for approximately 2 h, but not for the ACSF and mixture solutions.
(J, K) The colour change was obvious for three solutions after an overnight incubation for approximately 11 h under artificially controlled visible light irradiation.
(I) One milliliter of 50 mM DCIP was added again at 10:57 the next day (dark blue).
(M-P) The colour change was observed almost simultaneously in three reaction solutions after incubation for approximately 2 h, and the dark blue became colourless for both solutions containing ACSF.
Statistical results for Fig. 4

(A-C) Representative dynamic changes in biophotonic activity demonstrated by the RGVs are shown in the upper left panel. The sum of the time course of the average change in the RGVs from 30 continuously processed original grey images is shown in the upper right panel. One of two hemisphere slices (upper slice in the lower left regular photograph) was perfused independently with the recompetent glutamate treated with ~3 g freshly prepared chloroplasts for 48 h under light illumination after glutamate’s incompetent treatment with 4 whole brain slices for 24 h. Another slice (lower slice in the lower left regular photograph) was perfused independently with the incompetent treatment without chloroplast incubation. Representative biophoton grey images at the selected time periods indicated in the upper left panel and digit: 1-4 mark the 90, 270, 360, and 450 series of images, respectively (30 s imaging time for each image). The red, pink and black arrows indicate the time points for application of perfusion solution (0 min), washing (150 min) and reapplication (210 min), respectively. (A) A 160.25-370.70% increase in biophotonic activity was obtained for incompetent glutamate (n=5) compared to the reference induced by 50 mM p-Glu at the different imaging periods (n=37). (B), The biophotonic activity was increased for the ACSF treatment with ~3 g of freshly prepared chloroplasts after incubation with 4 whole brain slices for 24 h but were less than that in A (n=5). (C) ACSF treated with ~3 g of freshly prepared chloroplasts for 48 h resulted in increased biophotonic activity (n=5), which was reversed by adjusting the acidified ACSF to the original level of pH 7.4, indicating the acidification of ACSF by chloroplast.
Data are shown as the mean±s. e. m. n= the number of slices from the same number of mice. The reference data from 50 mM p-Glu in A (right panel) are the same samples as in Fig. 3C (right panel). Significant differences (*p<0.05 or ** p<0.01) are noted for A-C (see also Table S2).
Discussion
This study demonstrated that, after induction of neural biophotonic signals, glutamate lost its action and presented a decline in the molecular quantum energy levels, but it did not involve the changes in its molecular structure. Although the exact quantum biological mechanism, including the atoms involved, is unclear, our findings suggest that this phenomenon may be highly associated with the active hydrogen atom on the glutamate molecule. The change in the quantum states (quantum levels) in a liquid molecule involves the translational level of the molecule, the rotation, vibration and motion of the electrons, and the change in the energy level of the nuclear motion in the molecule (31-33). The mechanism of the decrease in the quantum energy levels after the action of glutamate on its receptors may be mainly related to the change in the electron quantum energy levels in the active hydrogen atom.
Therefore, our findings may provide a new idea for further clarifying the importance of the quantum biological mechanism of the functional molecules in the brain. For example, the glutamate released into the synaptic cleft is reused mostly by uptake into surrounding glial cells via the glutamate-glutamine (Glu-Gln) cycle, which is related to the synergism of neurons and glial cells, glutamate transporter and certain key enzymes (34, 35), and this widely-known process of recycling of glutamate seems to violate the energy conservation principle of the organism. Our findings suggest that such a process of recycling is indeed a necessary step for recovering the quantum energy levels of glutamate after its action, possibly as an important mechanism of quantum biology. Second, our findings could lead to a new perspective for elucidating the pathophysiological mechanisms of neuropsychiatric disorders. The changes in the glutamate recirculation, including the changes in transporters and key enzymes, are related to the pathological mechanisms of these diseases (14-21), while the action of an enzyme is considered to be a quantum mechanism, which is involved in the transfer of quantum energy (3). Therefore, the disruption of the Glu-Gln cycle may play an important role in the hypofunction of the glutamatergic pathway and contribute to the pathological mechanisms of neuropsychiatric disorders. Finally, our findings could provide new explanations for some clinical therapies such as local brain electromagnetic stimulations, electroshock, traditional natural medicine and acupuncture treatments (36, 37). One of the possible mechanisms for these treatments is through the restoration of the imbalance of the quantum energy levels of neurotransmitters and other functional molecules. In addition, the differences in the quantum energy levels of odorants may provide a novel way of explaining the diversity of the olfactory sense (2).
Methods
The detailed information for materials and methods is described in Supporting Information (SI).
Author contributions
J.D. conceived and designed the experiments; R.Z, T.C, and J.D. performed the experiments and analyzed the data; Z.W, and F.X. provided technical support for UBIS and research materials; J.D. wrote manuscript.
Competing financial interests
The authors declare no competing financial interests.
Acknowledgments
This work was supported by the Sci-Tech Support Plan of Hubei province (2014BEC086) and the research funds of South-Central University for Nationalities (XTZ15014 and CZP 18008), and partially by the National Natural Science Foundation of China (31640034, 31700911).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}