

Abstract
Multiplexed IF imaging using same species antibodies is in great demand as multiplexed IF is a general practice in biological and clinical laboratories whilst antibodies from different species are hardly available. We here presented a robust method to eliminate the use of secondary antibodies by replacing secondary antibody signal amplification with DNA-based Hybridization-Chain-Reaction signal amplification. We achieved as much as 89-fold signal enhancement, and four target imaging using all mouse primary antibodies.
Immunofluorescence (IF) imaging using antibodies is widely used in biological and clinical laboratories to map the distribution of proteins in situ. A typical IF protocol involves labeling specific targets with primary antibodies followed by signal amplification using secondary antibodies targeting the primary antibodies. This poses a significant challenge for multiplexed imaging as primary antibodies from different species or different subclasses (in the case of mouse antibodies) have to be used for the same sample. Unfortunately, most of the validated antibodies are produced either from mouse or from rabbit, hindering flexible antibody choices. Primary antibodies directly conjugated with fluorophores can be used to bypass the secondary antibody species issues. However, lack of signal amplification from secondary antibodies restricts this approach to the visualization of high abundance targets. In order to address the abovementioned challenges, we integrated a DNA-based signal amplification method, Hybridization Chain Reaction (HCR)1–4, with IF to achieve high quality multiplexed imaging using same species primary antibodies (Figure 1).

Cellular targets (T1, T2 and T3) are labeled with HCR-initiator (B1, B2 and B3) conjugated primary antibodies. HCR hairpins conjugated with fluorophores are then introduced to amplify the signals followed by visualization using conventional fluorescence microscopy.
In HCR, a single DNA initiator sequence will trigger an assembly of a large linear DNA structure by iterative HCR fluorescence hairpin opening1–4. We directly conjugated HCR-initiator DNA sequences to primary antibodies through covalent chemical modification5. We first tested the labeling specificity of DNA-modified primary antibodies by staining the antibodies in cultured BSC1 cells followed by HCR amplification. Surprisingly, we observed strong nonspecific signals, especially inside the nucleus (Figure 2 and Supplementary Figure 1), which has not been observed previously using DNA-conjugated secondary antibodies6,7. We decided to optimize the antibody staining protocol to reduce nonspecific binding. Sheared sperm DNA has previously been used to block nonspecific binding of DNA-conjugated antibodies8. We showed that addition of 0.5mg/ml of sheared sperm DNA during blocking and antibody incubation could reduce but not completely abolish nonspecific nuclear signal (Figure 2 and Supplementary Figure 1). In order to further suppress nonspecific signals, we tested the addition of dextran sulfate in the antibody incubation buffer. High concentration (10% - 20%) of dextran sulfate has been used as a crowding reagent in DNA and RNA FISH experiments9,10. Empirically, we noticed that some primary antibodies failed to bind to their targets in 10% dextran sulfate solution (data not shown). We therefore performed a titration experiment to determine the minimal concentration of dextran sulfate. The result showed a good suppression of nonspecific nuclear signal could be achieved with as low as 0.02% dextran sulfate (Supplementary Figure 2). We used 0.05% or 0.1% dextran sulfate plus 0.5mg/ml sheared sperm DNA for all following experiments. Similar to conventional immunostaining, we also noticed for some antibodies the antibody concentration also had a modest effect on the nonspecific signal (Supplementary Figure 1).

(a) BSC1 cells were fixed and stained with HCR initiator (B3)-conjugated Golgi 97 antibodies. The antibodies were diluted in buffers with/without 0.5mg/ml sheared sperm DNA and with/without 0.05% dextran sulfate. Antibodies were diluted using 1:50 or 1:200 dilution factor. Scale bar: 20 μm. (b) Quantification of nuclear signal. Error bar is SEM and n = 100.

BSC1 cells are fixed and stained with HCR initiator DNA (B1 or B5) conjugated primary alphaTubulin and betaTubulin antibodies. Different concentration of dextran sulfate (0% to 10%) in the antibody incubation buffer was tested. (a) Demonstration of nonspecific nucleus signal in alphaTubulin-B1 and betaTubulin-B1 images. Scale bar: 10 um. (b) Quantification of nonspecific nuclear signals. (Error bar is SEM. N = 30)

(a) BSC1 cells were fixed and stained with HCR initiator (B1 or B5)-conjugated antibodies against either alphaTubulin or betaTubulin. The antibodies were diluted in buffers with/without 0.5mg/ml sheared sperm DNA and with/without 0.1% dextran sulfate. AlphaTubulin images were displayed at grayscale 0-255, and betaTubulin images were displayed at grayscale 0-100. Scale bar: 10 μm. (b) Quantification of nuclear signal. Error bar is SEM and n = 30.
We then quantified the signal amplified using HCR compared with the unamplified signal and the signal amplified using commercial fluorophore-conjugated secondary antibodies. We stained cultured BSC1 cells or primary neurons with HCR initiator B1-conjugated alpha-tubulin antibodies, B5-conjugated alpha-tubulin or B5-conjugated GFAP antibodies (Figure 3). We were able to achieve as much as 89-fold increase of fluorescence signal for alpha-tubulin using HCR (Figure 3). More importantly, HCR outperformed commercial fluorophore-conjugated secondary antibodies and achieved better signal amplification.
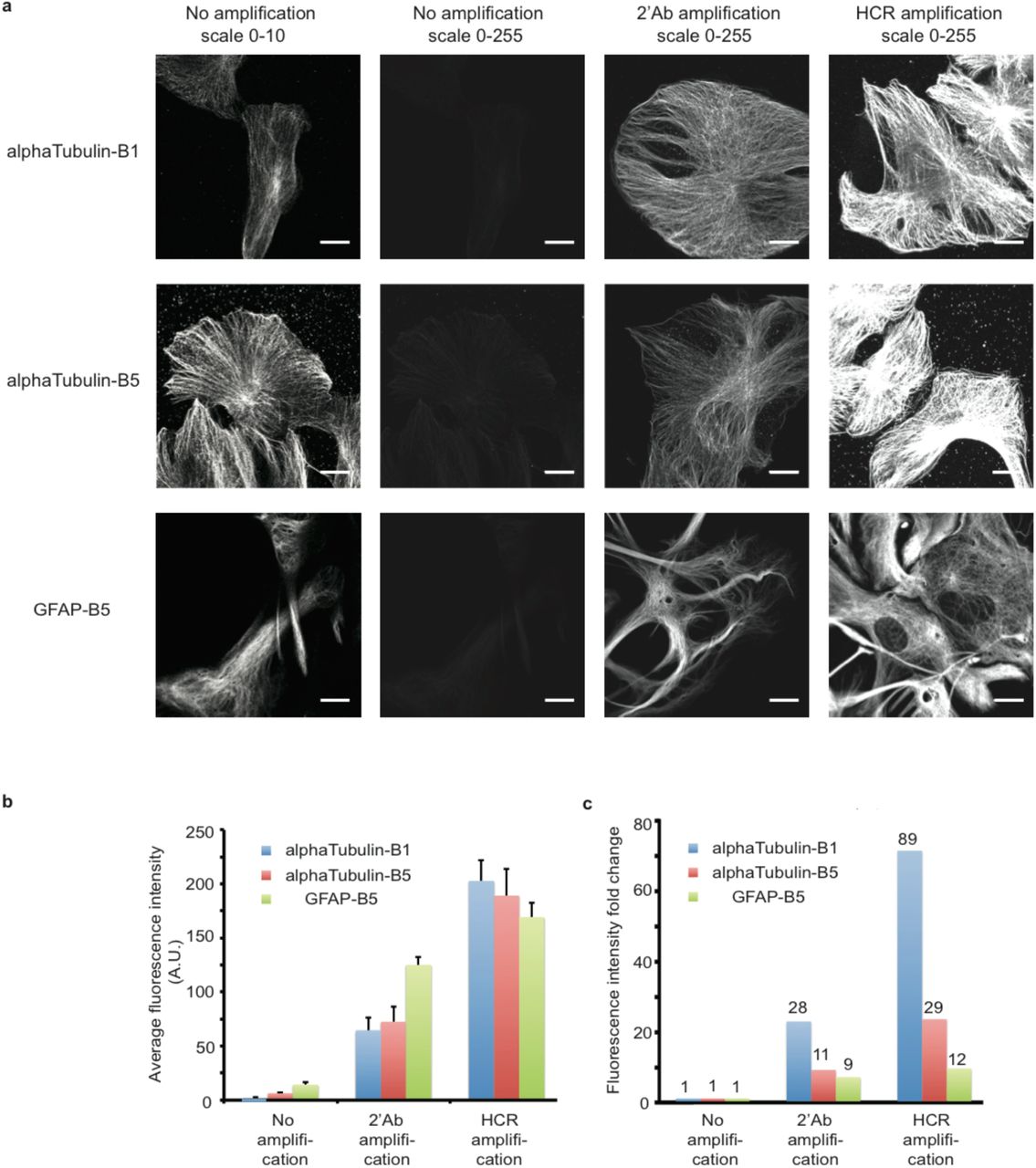
Quantification of fluorescence signal amplification by HCR.
(a) BSC1 cells or primary mouse hippocampal neuron culture were fixed and stained with HCR initiator-conjugated antibodies against alphaTubulin and GFAP, respectively. In no amplification samples, the images were acquired using Alexa647-conjugated DNA strands that are complementary to the HCR initiator DNA sequences. Either Alexa647-conjugated secondary antibodies (2’Ab) or HCR hairpins were used to amplify the signals. The images were acquired at the same microscope setting. Scale bar: 20 μm. (b) Quantification of image fluorescence intensity. Error bar is SEM and n = 12. (c) Quantification of fluorescence intensity fold change based on average fluorescence intensity in b).
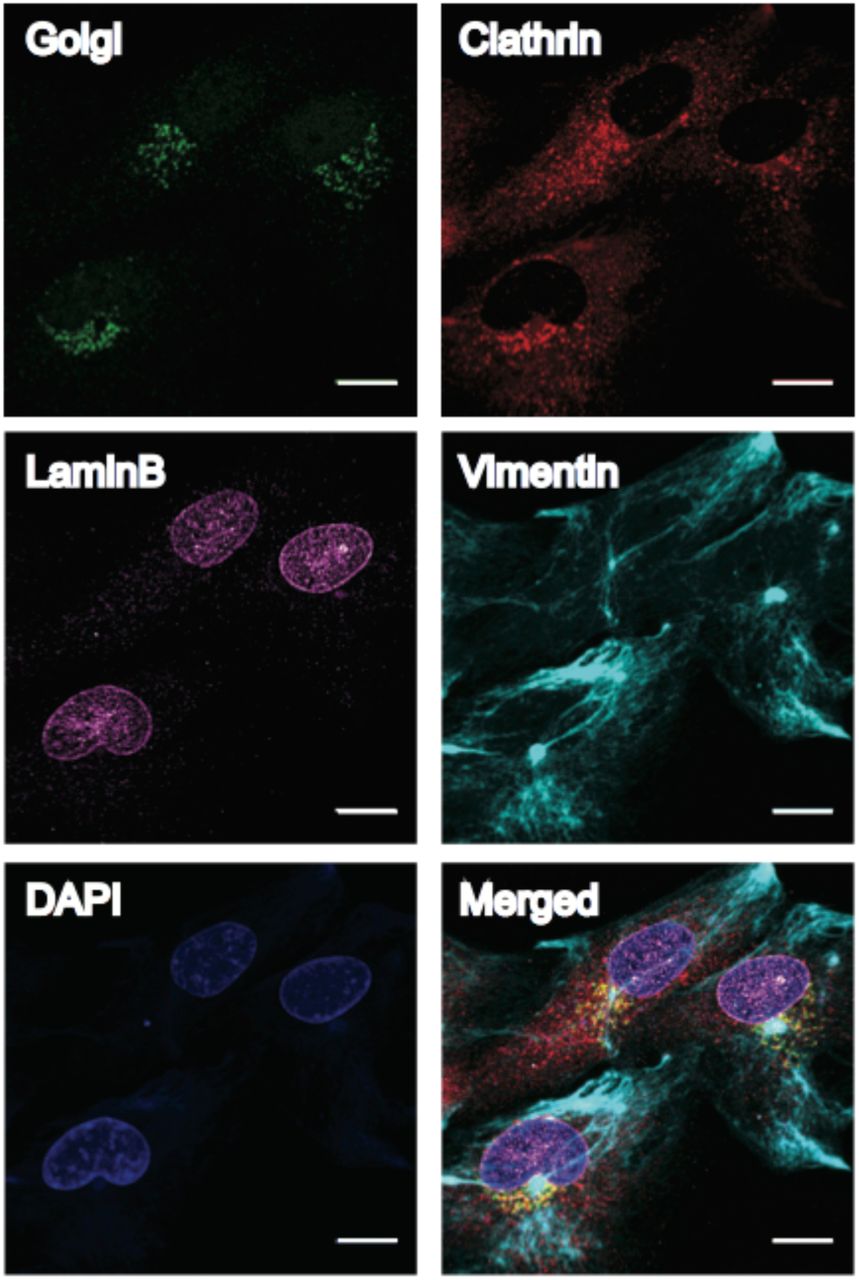
We finally performed multiplexed imaging using same species primary antibodies by conjugating different HCR initiator sequences to different antibodies. We first attached four mouse IgG (IgG1 subclass) primary antibodies against four distinct cellular structures (Golgi 97, Lamin B, Vimentin and Clathrin) with HCR initiators (B2 to B5). We then stained cultured BSC1 cells with the four antibodies using our optimized staining protocol, followed by HCR signal amplification and confocal microscopy imaging. We were able to get high-quality images for all four targets (Figure 4). We also performed multiplexed imaging in mouse retina tissue samples using four mouse IgG primary antibodies targeting Bassoon, SV2, pNFH (phosphorylated neurofilament heavy) and Calretinin. Bassoon and SV2 are two synaptic markers, and pNFH and Calretinin are two neuronal markers. Bassoon is located in the active zone of synapses, and is in low abundance11. It can be seen that after HCR signal amplification, Bassoon signals are clearly visible in 40 μm thick mouse retina sections (Figure 5).

Four mouse IgG1 antibodies are conjugated with HCR-initiators (Golgi-B3, Clathrin-B2, LaminB-B4 and Vimentin-B5) and used to stain fixed BSC1 cells. DAPI was used to visualize the nucleus. Scale bar: 20 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
40 μm mouse retina sections were stained with HCR initiator-conjugated primary antibodies against targets as labeled (Bassoon-B1, SV2-B2, pNFH-B4 and Calretinin-B5). (a) Visualization of synapses marked by Bassoon with 63x microscope magnification. Left panel scale bar: 50 μm; Right panel scale bar: 20 μm. (b) Five-target imaging (four protein markers plus DAPI) visualized with 20x microscope magnification. Image stacks were taken and projected using a maximum projection algorithm. Scale bar: 50 μm.
Multiplexed IF imaging using same species antibodies is in great demand as multiplexed IF is a general practice in biological and clinical laboratories whilst antibodies from different species are hardly available. We here presented a robust method to eliminate the use of secondary antibodies by replacing secondary antibody signal amplification with DNA-based HCR signal amplification. A general concern regarding conjugating DNA to antibodies is that the attached DNA can affect antibody binding affinity and specificity. We have optimized the antibody staining protocol to reduce nonspecific antibody binding. In addition, our current DNA conjugation protocol has been optimized to attach only ∼1.5 DNA oligonucleotides per antibody to minimize the chance disrupting epitope recognizing regions5. Site-specific conjugation methods, such as conjugation to the Fc domain of antibodies through protein G12, have also been reported and can be used to further reduce the possibility of affinity loss. Although only five targets imaging (four protein targets plus DAPI) is demonstrated here, higher multiplexing is achievable by developing more orthogonal HCR pairs, which is currently under development.
Author contribution
Y. W and G.M.C conceived the project. Y.W. designed and performed the experiments. W.X performed the experiments. R.E.K and G.M.C. supervised the project. Y.W. wrote the manuscript.
Acknowledgements
We thank Dr. Demian Park for providing primary mouse hippocampal neuron cultures. We thank Dr. Sylvain Lapan for providing mouse retina samples. This work is supported by NIH grant (RM1HG008525) and IARPA grant (IARPA MICrONS; D16PC0008).




