

Abstract
Disturbed sleep homeostatic states have been found to alter neuronal homeostasis and reduce grey matter (GM) volume. Caffeine intake that interferes with sleep homeostasis through antagonizing adenosine receptors can impair hippocampal synaptic strength, neurogenesis, as well as memory and learning in rats. In this study, reduced medial temporal GM volume was observed after daily caffeine intake in humans (3 × 150 mg × 10 days compared to 10-day placebo administration). The potential bias from reduced cerebral blood flow was controlled, and the GM reduction was independent of the change in sleep pressure. A decrease in working memory during daily caffeine intake was observed, albeit no association with the magnitude of GM changes. The findings indicate that daily caffeine intake might induce rapid cerebral plasticity and be detrimental for higher order cognitive performance in the long run. They may call into question whether the neuroprotective effects of caffeine found in acute or low dose administration in animals are generalizable onto the daily usage in humans.
Introduction
Caffeine is the most commonly used psychostimulant worldwide and mainly consumed in forms of coffee, tea, energy drink, and soda 1–4. Although caffeine is mostly considered to be non-addictive, the observed physical and psychological dependence 5,6 consolidate its regular consumption 7–9 through the caffeine-induced reinforcing effects 10, as well as the motive to resist withdrawal symptoms 11 and to increase alertness 12. Higher alertness after acute caffeine intake 13 mirrors a reduced homeostatic sleep pressure, which is also evident in a reduced depth of sleep 14. The latter is characterized by attenuated electroencephalographic slow-wave activity (EEG SWA, 0.75 – 4.5 Hz) in non-rapid eye movement (NREM) sleep and shortened slow-wave sleep (SWS) 14–17.
Disturbed sleep homeostasis can not only lead to cerebral micromorphometric alterations in the mitochondria and chromatin that leads to cell death 18–20, but also macrostructural changes. Lower GM volumes were observed during abnormally high sleep pressure, such as during sleep deprivation or sleep fragmentation. Liu, et al. 21 reported reduced thalamic GM volume along with impaired cognitive performance in healthy adults after 72-hour prolonged waking compared to baseline. Dai, et al. 22 demonstrated a GM dynamic through the 36-hour course of sleep deprivation, where a decrease in right thalamus, right insula, right inferior parietal lobe, and bilateral somatosensory association cortex was observed by 32 hours of sleep deprivation. In clinical studies, GM volume and cortical thickness were reduced in patients with various sleep disorders (e.g. chronic insomnia 23,24, sleep apnea 25, and narcolepsy 26) compared to healthy controls. Furthermore, the higher symptomatic severity of insomnia and narcolepsy was commonly associated with the reduced frontal GM. The micro- and macro-morphometric changes in GM in response to sleep deprivation might reflect the disrupted adenosine-modulated cellular homeostasis, such as cardiac microtubule dynamic 27, astrocytic cytoskeleton arrangement 28, hippocampal fiber synaptic plasticity 29, and the robustness of cortical axons and dendrites 30,31.
Caffeine interferes with sleep homeostasis by dampening the accumulation of sleep pressure through the antagonism on adenosine A1 and A2A receptors 15,32. Evidence in animals shows that acute or long-term caffeine consumption inhibits the long-term potentiation (LTP) 33, neurogenesis 34,35, and cell proliferation 36 in hippocampus, and can impair learning and memory 34. However, it is still not clear whether daily caffeine consumption leads to cerebral structural modifications in healthy populations through the constant impact on sleep homeostasis, despite a number of studies exploring the functional neuroprotective effect of caffeine on Parkinson’s and Alzheimer’s disease 37,38.
Hence, we hypothesized that, through changes in sleep homeostasis, long-term caffeine intake alters GM structures, and later, we tested exploratorily whether the GM alterations were related to changes in memory function. Cerebral GM volume was measured by magnetic resonance imaging (MRI) together with cerebral blood flow to control for its bias on MRI signals, and sleep pressure was indexed by nighttime sleep EEG slow-wave activity. Memory performance was assessed by verbal N-back tasks.
Results
Caffeine-induced reductions in total GM volume and in medial temporal regions
Total GM volume was lower in the caffeine condition compared to placebo (tcon = −3.59, 95% CI= [−0.009, −0.003], p < .001, Figure 1). A voxel-based analysis (VBA) indicated that the reduction of GM volume was most prominent in the right medial temporal lobe (mTL, including hippocampus; VBA cluster-level pFWE = .032, Figure 2a; eigenvalue tcon = −4.35, 95% CI = [−0.012, −0.005], p < .001), along with the left frontal pole, right postcentral gyrus, right insula, and the cerebellum at trend (i.e. VBA pFWE < .063, pFDR = .029). No increases of GM in the caffeine compared to the placebo condition were observed.
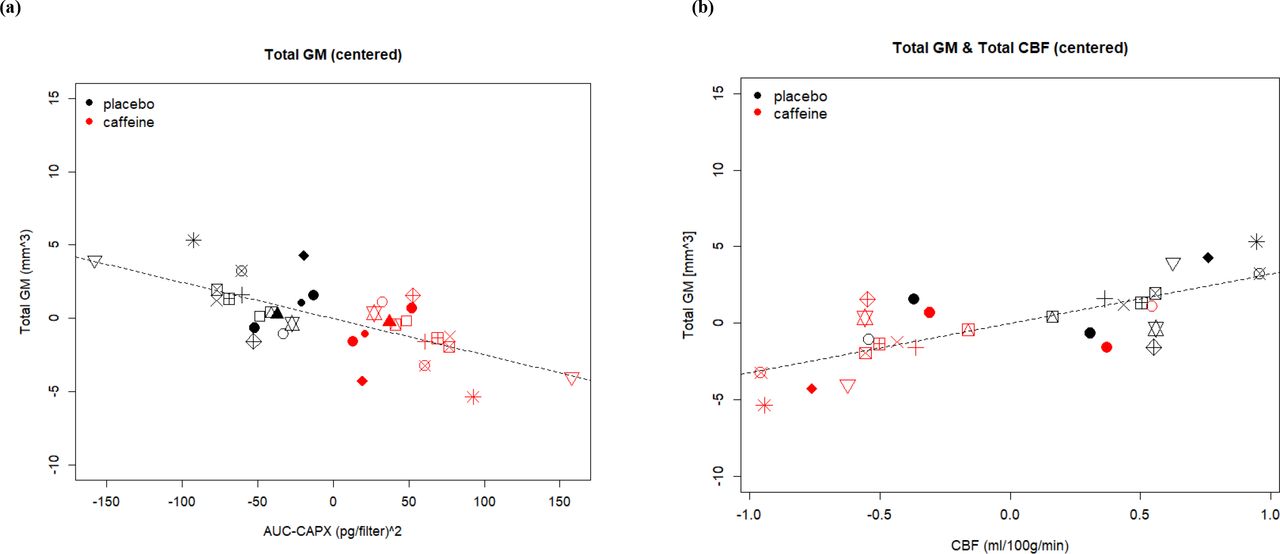

We use center a plot to display the changes in a specific variable to the treatment of caffeine and placebo in each subject. The values are the relative distance from the responses in each condition to the average response of a single participant, calculated as responsecaff or plac – (responsecaff + responseplac)/2. Each symbol represents one participant. In two dimensions one can observe the variance of within-subject changes of two variables between conditions (by color), as well as observe the association between the changes of two variables (x and y axes). The stronger the condition effect is, the more discretely are the two-colored clouds distributed. The stronger the association between two variables is, the closer the shape is to linear. (a) and (b) Total GM volume is associated with AUC-CAPX negatively and with total CBF. (c) Between-subject variance of Total GM volume is associated with NREM SWA especially in placebo condition (presented in raw data points). Reasons for missing data points are addressed in supplement.


Voxel-based and region of interest (ROI)-based approaches were both employed. The dual process allows a better spatial accuracy when first determining the condition effect on GM, while the extracted eigenvariate provides further modeling with other variables (AUC-CAPX, SWA, and CBF) to reduce bias caused by multiple comparison correction. (a) Voxel-based analysis revealed a significant GM reduction in mTL in the caffeine condition compared to placebo (color bar displayed in pFWE). (b) The individual responses in GM volume within the significant mTL clusters was positively associated with the levels of caffeine and paraxanthine. (c) No significant association between mTL GM and mTL CBF was found. Note that there the missing data points and reasons are addressed in supplement.
For further analyses, we extracted the eigenvariate of the significant cluster in the right medial temporal lobe, to which we will refer to as mTL GM volume. In order to support that the observed changes in mTL GM are induced by our treatment rather than related to a confounder, the association between the individual GM differences and the levels of caffeine and paraxanthine (AUC-CAPX) was examined (for AUC-CAPX levels during each condition across time see supplement). A negative association between AUC-CAPX and both total GM (tAUC = −3.46, 95% CI = [−0.00008, −0.00002], pAUC = .001; Figure 1a) and mTL GM (tAUC = −3.83, 95% CI = [−0.004, −0.001], pAUC = .001; Figure 2b) was found, namely the higher the AUC-CAPX the lower was the GM volume . The overview of the results and effects of coefficients are reported in Table 1.
The pairwise association and its coefficient of the effects between all physiological variables. The superscripts indicate the corresponding models stated in the supplement. The coefficients of the effects (R2) on each model were calculated during the linear mixed model estimation, including the R2 marginal (R2m) for the coefficient of fixed effect and the R2 condition (R2c) for the coefficient for fixed + random effects.
Caffeine-induced differences in the association of SWA and GM volumetric reductions
In a next step, it was examined whether SWA-indexed sleep pressure is reduced during long-term caffeine intake compared to placebo, and whether the changes in GM were mediated by SWA. Since SWA in the first NREM episode shows a high sensitivity to acute caffeine intake 17,39, this specific segment of the night was adopted to index homeostatic sleep-pressure in our study. The overview of the results and effects of coefficients are reported in Table 1.
No significant change in NREM SWA after 9 days of daily caffeine intake compared to 9 days of daily placebo intake was observed (tcon = −0.87, pcon = 0.386, see supplement). Yet, overall SWA was positively associated with total GM (tSWA = 2.88, 95% CI = [0.00008, 0.0004], pSWA = .004, Figure 1c), and a significant interaction between condition and SWA indicated a stronger association in the placebo condition (tinter = −4.709, 95% CI = [−0.0002, −0.0001], pinter < .001). In the regional mTL GM differences, no associations between SWA and GM volumetric reductions were seen (tSWA = 0.09, pSWA = .931).
CBF response during daily caffeine consumption
When estimating GM tissue volume, caffeine-induced reductions in regional CBF can regionally bias the MRI signal distribution and therefore the tissue probability maps 40–42. Thus, we first examined the CBF response during caffeine intake compared to placebo, followed by the inspection of its contribution to the observed GM changes. The overview of the results and effects of coefficients are reported in Table 1.
A significant reduction in total CBF was found in the caffeine compared to the placebo condition (tcon = −5.2, 95% CI = [−0.119, −0.054], pcon < .001). Voxel-wise analysis revealed that reductions occurred mainly in the midline, including cuneus, precuneus, and subcortical regions (pFWE_all < .05, Figure 3). Importantly, also in the region where a caffeine-associated GM volumetric reduction was observed, CBF was also lower in the caffeine condition compared to placebo (tcon = −2.17, 95% CI = [−2.644, −0.132], pcon = .048). AUC-CAPX was negatively associated with both total CBF (tAUC = −6.33, 95% CI = [−0.0009, −0.0005], pAUC = .001) and mTL CBF (tAUC = −2.77, 95% CI = [−0.021, −0.004], pAUC = .055).

Regions (in red) showing a reduction in CBF after caffeine intake. Most prominent reduction were observed in cuneus, precuneus, and subcortical regions (pFWE < .05).
Despite a reduction in both total and mTL CBF, the associations with GM volume were different. A positive association between the reduction of total CBF and total GM volume was observed (tCBF = 4.70, 95% CI = [0.004, 0.009], pCBF < .001). Moreover, the variance of CBF mediated caffeine-associated reductions on total GM: Including total CBF as a covariate fully accounted for the main effect of condition on total GM estimation (tCBF = 2.82, pCBF = .005; tcon = −0.77, pcon = .441, Figure 1b). In contrast, CBF did not account for the caffeine-induced volumetric reductions in mTL GM, as indicated by multi-modal VBA with linear mixed model. Consistently, in the ROI-based analysis, no association between the caffeine-associated changes in mTL CBF and in mTL GM was found (tCBF = −1.67, pCBF = .122, Figure 2c). Thus, reasonably, no mediation was exerted by mTL CBF on the mTL GM volumetric reduction (tCBF = −1.88, pCBF = .083; tcon = −4.80, pcon < .001).
Working memory performance during daily caffeine intake
In a last step, a possible functional implication of the caffeine-induced mTL GM reductions was explored. To do so, performance in a verbal n-back task with two levels of difficulty (0-back, referred to as low load; 3-back, referred to as high load) was analyzed with adjustment for the order of the conditions. The task was administered every 4 hours during 12.5 hours before and during the MRI measurements. The differences in average accuracy and reaction time (RT) of four attempts between the caffeine and placebo condition were examined. The overview of the results, 95% CI, and effects of coefficients are reported in the supplement.
Compared to placebo, lower accuracy was found in both 0-back (tcon = −2.33, pcon = .020, Figure 4) and 3-back (tcon = −2.02, pcon = .044, Figure 4) performance in the caffeine condition, as well as a lower net accuracy of 3-back, i.e. corrected for the baseline response of 0-back performance (tcon = −1.97, 95% CI = [−0.0355, −0.00010], pcon = .049; R2m = .41, R2c = .85). The reductions in accuracy were not linked to the reductions in total or regional GM in caffeine compared to placebo. Moreover, in contrast to the worse accuracy in the caffeine condition compared to placebo, a higher AUC-CAPX level within the caffeine condition was positively associated with a better net 3-back accuracy (tAUC = 3.58, pAUC < .001). No significant association of AUC-CAPX and 0-back accuracy was observed. On the other hand, while there was no difference found in the net overall RT and the RT of hits and false alarm responses between two conditions (tall < 1.88, pall > .60), the RT of missed (tcon = 2.75, pcon = .006) and correction rejections (tcon =1.95, pcon = .051) were longer in caffeine condition compared to placebo. The net RT in all types of responses, however, was negatively associated with AUC-CAPX (tall < −2.44, pall < .015).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In the caffeine condition, the accuracy was significantly worse in both 0-back and 3-back compared to placebo. We observed no significant difference in overall RTs.
Discussion
Our study examined whether daily caffeine intake alters cerebral morphometry, and whether these changes are mediated by homeostatic sleep pressure, as indexed by sleep EEG slow-wave activity (SWA). We observed reductions in total grey matter (GM) volume, which were accounted for by cerebral blood flow (CBF), suggesting a strong necessity to strictly control for both caffeine intake and CBF in future studies of GM volume. However, we also observed caffeine-induced concentration-dependent reductions in GM volume in a cluster within the medial temporal lobe (including hippocampus, parahippocampus, fusiform gyrus), which were not accounted for by the reduction in CBF. In contrast to our hypothesis, daily caffeine intake did not reduce EEG slow-wave activity during the night sleep, nor were the GM reductions directly mediated by alterations of sleep SWA. Caffeine might, thus, exerts an effect on cerebral plasticity through a parallel pathway to its influence on sleep, as discussed in the following sections. In a nutshell, our data indicate that daily intake of caffeine, as observed in around 80% of the population in the world 43, challenges GM plasticity. Together with the observed impairments of working memory performance, they strongly question earlier-reported benefits of daily caffeine intake especially in healthy populations.
A caffeine-induced lower GM volume may reflect multiple possible responses, one main assumption is neuronal damage. It has been observed that daily or high-dose caffeine intake weakens A2AR affinity for caffeine 44 and lead to an upregulated A2AR availability 45,46, consequently maintains boosting in glutamate signaling and prolonged post firing of hippocampal pyramidal cells 44,47,48. Furthermore, the strengthened A2AR agonism decreases the affinity of A1R for caffeine on the A1-A2A receptor heteromers, which is speculated to result in a tolerance effect in behaviors after daily use of caffeine 44,49 and, potentially as observed in our study, SWA.
Sleep SWA is positively linked with adenosine binding and dissipation during sleep 50. SWA renormalizes the saturated synaptic capacity and recover the brain neurons from the energy consumption during prior wakefulness, suggested by the synaptic homeostasis hypothesis (SHY) 51. The strength of synaptic potentiation can be regained only if the renormalization of synaptic strength has occurred during sleep 52. Our data suggest that chronic caffeine intake might affect this homeostatic balance. We observed a between-subject concentration-dependent enhancement on memory performance that might reflect the boosting effect of caffeine on cerebral activities. At the same time, however, there was lack of a commensurate response in SWA as the due synaptic recovery from the toned-up neuronal strength. A deficient synaptic restoration followed by the continuous administration of caffeine on the next day may presumably maintain the cellular stress, inhibit the functions of hippocampal pyramidal cells 33–35 due to exhaustion, and, in the long run, impair memory performance as observed in our study as well as other animal studies on daily administration of caffeine 34 and adenosine antagonists 53.
Beside a change in neurons, the lower GM observed in our study can also result from changes in non-neuronal cells and/or in cerebral vasculature 54. In oncological studies, caffeine has been applied to induce apoptosis of glial cells 55. Despite an absence of direct evidence on the effect of daily caffeine intake, the effects of adenosine and its A2A receptors on the release of growth factors have been strongly suggested 38, which in turn modulate the proliferation of astrocytes and can influence angiogenesis 56.
Changes in GM volume, as derived by VBM, can also be simply confounded by differences in CBF 40–42. Accordingly, in the present study, CBF accounted for total GM changes induced by caffeine intake. These apparent total GM changes emphasize the importance of controlling for caffeine consumption in repeated fMRI measures, especially with a region of interests in the most impacted regions (cuneus, precuneus, cerebellum, and subcortices). The observation of the typical caffeine-induced reductions in CBF 42,57,58 in our study, moreover add to the current knowledge that the history of caffeine intake at least within 4.5 hours strongly impacts on both CBF and apparent tissue change.
Our treatment did not elicit a significant effect on SWA in the first NREM sleep episode. The contradiction to earlier findings14, 16,17 can be reconciled within two assumptions: a timing-dependent influence of caffeine on sleep, and a potential development of tolerance in response to the repeated daily intake. Earlier studies indicate that caffeine exerts strong reduction in NREM SWA when caffeine was administered close to bedtime 16,17 while the effects were considerably weaker after morning administration 59. Moreover, a tolerance effect to caffeine treatment was found in other sleep features during a 11-day course 60, which might similarly apply to SWA.
The combination of presence and absence of caffeine effects on CBF and SWA respectively further suggest distinct responses of A1R and A2AR in developing tolerance to daily intake 46,61–64. While the antagonism on A2A and A2B receptors plays a predominant role on vasoconstriction 65,66, the sleep homeostatic regulation is primarily modulated by A1R through the cholinergic signaling in basal forebrain 14,15,67 together with an indirect modulation from A2AR 68. Therefore, while daily intake of caffeine reduces CBF through the blockade of A2AR without developing complete tolerance, the tolerance response of A1R might result in the divergent response in SWA. Furthermore, chronic caffeine treatment is found to strengthen A2AR agonism on its A1R-heteromers, which leads to a decrease in caffeine binding to A1R and speculatively result in a tolerance effect in behaviors 44,49, and potentially, SWA.
Notably, we also observed a positive between-subject association between SWA and total GM volume particularly in the placebo condition, which might reflect the individual variance of brain maturity during young adulthood 69. However, the attenuated association in caffeine condition might simply be caused by the increased variance due to the bias of CBF after caffeine intake. On the contrary, GM in the mTL region sensitive to our treatment did not exhibit this association, which potentially reflects region-specific mechanisms of plasticity during sleep 70.
As the first study about caffeine effects on GM in young healthy adults, our results may seem contradictory to earlier evidence on its neuroprotective effect on cognitive impairment 71,72. However, studies that demonstrated the neuroprotective effects of caffeine/adenosine antagonists were mostly based on preventive models, where one observed the efficacy of the agent in reversing neurodegeneration or inflammation induced by kinase, transgenicity, or trauma 8,73,74. The effects therefore cannot be generalized and thus have left the puzzles on how the constant interference of caffeine on a healthy adenosinergic system would alter the intrinsic neurotransmission. Moreover, there is a considerable inconsistency in the observed neuroprotective effects of caffeine and other adenosine antagonists75. For instance, a high-dose administration of A2AR antagonists can induce excitotoxicity acutely in vivo and ex vivo 76,77. A peripheral administration and a direct neuronal injection of identical selective A2A antagonist might also lead to opposite outcomes on the neuroprotective effects 78,79. A recent study suggested that age-related cognitive and neuronal declines were ameliorated by chronic caffeine consumption, yet also with lower dose (5 mg/kg/day) as compared to our study 80. Given the distinct responses and interaction between the subtypes of adenosine receptors, the difference between pre- and post-synaptic administration, and the dose-dependent effect, it is of importance to carefully compare the study designs that focused on a similar research question.
Our study also bears some limitations that require a careful interpretation. Firstly, although a power calculation indicated a sufficient sample size, we came across data loss due to technical reasons, which lowers the signal-to-noise ratio to detect a relatively small effect. To address this issue, a non-parametric permutation test was therefore employed in all the voxel-based analyses as an initial step, and the subsequent analyses were established upon its results. Secondly, one might argue that the absence of a significant difference in SWA might be due to a genetically predisposed insensitivity to caffeine of the selected population 81. However, as the withdrawal from daily intake of 450 mg of caffeine has been shown to induce clear-cut responses in the sleep-homeostatic regulation of these participants 82, it seems unlikely that the selected population was insensitive to the stimulant. Notably, the habitual amount was calculated from all types of caffeinated diets, not only coffee intake but also tea, chocolate drinks, energy drinks, soda, and so on.
Overall, our findings derived from this first within-subject laboratory study yield an insight on the mTL GM plasticity induced by the repeated intake of caffeine in a long-term course. Reductions in GM volume and the cooccurring impairment in working memory performance underline the importance of carefully questioning the consequences of habitual daily intake of a psychostimulant, freely available all over the world.
Acknowledgement
We sincerely appreciate the contribution of our interns Andrea Schumacher, Laura Tincknell, M.Sc. student Sven Leach, and all the study helpers assisting in the experiment. We also thank Dr. med. Corrado Garbazza and Dr. med. Helen Slawik for the health check during screening process. We acknowledge gratefully the collaboration with Prof. Christopher Gerner and Dr. Samuel Meier at University of Vienna, together with the assistance of M.Sc. Max Feuerstein, and Dr. Rupert Mayer for the measurement of fingertip sweats. We especially appreciate all our participants for their volunteering and cooperation. This project is financed by Swiss National Science Foundation (320030-163058).
References




