

ABSTRACT
Lanthanides are elements that have been recently recognized as “new life metals” in bacterial metabolism, yet much remains unknown regarding lanthanide acquisition, regulation, and use. The lanthanide-dependent methanol dehydrogenase XoxF produces formaldehyde, which is lethal to Methylorubrum extorquens AM1 if allowed to accumulate. This property enabled a transposon mutagenesis study to expand knowledge of the lanthanide-dependent metabolic network. Mutant strains were reconstructed and growth studies were conducted for over 40 strains detailing the involvement of 8 novel genes in lanthanide-dependent and independent methanol growth, including a fused ABC-transporter, aminopeptidase, LysR-type transcriptional regulator, putative homospermidine synthase, mxaD homolog (xoxD), porin family protein, and genes of unknown function previously published as orf6 and orf7. Using genetic and biochemical analyses, strains lacking individual genes in the lanthanide transport cluster were characterized and named lut for lanthanide utilization and transport (META1_1778 to META1_1787). Consistent with previous reports, we corroborated that a TonB-ABC transport system is required for lanthanide transport to the cytoplasm. However, an additional outer membrane transport mechanism became apparent after longer growth incubations. Additionally, suppressor mutations that rescued growth of the ABC-transporter mutants were identified. Transcriptional reporter fusions were used to show that like iron transport, expression from the TonB-dependent receptor promoter, lutH, is repressed when lanthanides are in excess. Energy dispersive X-ray spectroscopy analysis was used to visualize the localization of lanthanum in wild-type and TonB-ABC transport mutant strains and showed for the first time, that M. extorquens AM1 stores cytoplasmic lanthanides in mineral form.
IMPORTANCE Understanding the role of lanthanides in bacterial systems is an emergent field of study. Results from this work define mechanisms by which the methylotrophic bacterium, M. extorquens AM1, acquires and accumulates lanthanides. Lanthanides are critical metals for modern technologies and medical applications, yet lanthanide mining is cost-prohibitive and harmful to the environment. Methylotrophs are an attractive platform for the recovery of lanthanides from discarded electronics, mining leachate, and other waste streams as these microorganisms have been proven to sense and transport these metals for use in alcohol oxidation. Further, methylotrophs are effective biotechnological platforms for chemical productions and can use pollutants such as methane, and inexpensive feedstocks such as methanol. Defining the lanthanide acquisition, transport, and accumulation machinery is a step forward in designing a sustainable, clean platform to recover lanthanides in an efficient and less environmentally destructive manner.
INTRODUCTION
Lanthanide metals (Ln) impact the metabolism of many Gram-negative methylotrophic bacteria at both catalytic and transcriptional levels (1–3). Diverse types of pyrroloquinoline quinone (PQQ)-containing alcohol dehydrogenases (ADHs), specifically from the XoxF- and ExaF-families, have been shown to coordinate Ln in the active site (4–9) where they act as potent Lewis acids to facilitate a hydride transfer from the alcohol to PQQ, prompting alcohol oxidation (7, 10, 11). Some XoxF-methanol dehydrogenases (MeDHs) catalyze sequential oxidations to produce formate from methanol (12, 13), while formaldehyde is the product of other XoxF-MeDHs (8). Though much progress has been made regarding the catalytic role of Ln in ADH reactions, relatively little is known about how these Ln are acquired and incorporated into the enzymes that use them, and if they are stored intracellularly.
The role of Ln in metabolism is not exclusively catalytic. When Ln are transported into the cell, a transcriptional response occurs; this effect is commonly referred to as “the Ln-switch” or “rare earth-switch” (14–17). During the Ln-switch, the mxa operon is downregulated and transcript levels of the xox operon are upregulated (8, 15–19). In M. extorquens strains AM1 and PA1, the MxbDM two-component system and xoxF have been shown to be required for expression of the mxa genes and repression of the xox genes in the absence of Ln (20–22). However, suppressor mutations in the mxbD sensor kinase can arise which bypass the need for XoxF, presumably by constitutively activating the MxbM response regulator (3, 21, 22).
The machinery necessary for Ln transport is in the early stages of characterization and is predicted to be analogous to siderophore-mediated iron transport (22, 23). In M. extorquens AM1, ten genes predicted to be involved in Ln transport and utilization are clustered together in the genome (META1_1778 – META1_1787) and encode a TonB-dependent receptor, an ABC-type transporter, four exported proteins of unknown function, lanmodulin which binds Ln, and an additional periplasmic-binding protein capable of Ln binding (META1_1781) (23, 24). TonB-dependent receptors are commonly used for the transport of metal ions such as iron (Fe3+) and nickel (Ni2+) that form poorly soluble salts under physiological conditions (25, 26). To facilitate the uptake of insoluble Fe3+, these receptors have high affinity for metal-siderophore complexes. Siderophores are small molecules (hydroxamates, catecholates, and carboxylates) that are excreted by many bacteria and fungi to chelate insoluble metals (27–29). TonB is anchored in the inner membrane and spans the periplasmic space to interact with TonB-dependent receptors in the outer membrane. Once the siderophore-Fe3+ complex binds to the TonB-dependent receptor, it is transported through the receptor into the periplasm using the proton motive force harnessed by the TonB-ExbB-ExbD energy transducing system (30–32). In the periplasm, the siderophore-Fe3+ complex binds to a periplasmic binding protein that facilitates its interaction with an ABC-type transporter to translocate the siderophore-bound Fe3+ to the cytoplasm (33, 34). Iron release from the siderophore complex involves a reduction of Fe3+ to Fe2+.
In M. extorquens strain PA1, genetic studies disrupting either the TonB-dependent receptor or deletions spanning multiple genes in the Ln transport cluster showed the requirement of some but not all genes in this cluster for Ln-dependent methanol growth (22). Accordingly, Mattocks et al. identified homologs of this system in M. extorquens AM1 and showed that a putative exported protein encoded by META1_1781 can efficiently bind Ln from lanthanum (La3+) to gadolinium. Additionally, they showed that the periplasmic binding component of the ABC transporter encoded by META1_1778 was unable to bind Ln (23), possibly because it instead binds the predicted siderophore-like molecule necessary to transport Ln into the cell.
Currently, the regulation of the Ln-transport cluster genes is not understood. Iron transport is highly regulated as Fe2+ excess can promote Fenton chemistry and lead to the production of highly reactive radicals, which damage DNA, proteins, and other biomolecules (35). In bacteria such as Escherichia coli, the ferric uptake regulator (Fur) and the small RNA, rhyB, help to mediate iron homeostasis. When Fe2+ accumulates, Fur binds to iron uptake operons including fecABCDE (36), fhuACDB (37), cirA (38), tonB (39) and exbB-exbD (37) to repress iron transport and prevent oxidative stress. Herein, we show that like the iron paradigm, expression from the Ln TonB-dependent receptor is repressed by Ln when they are in excess.
In this study, we describe new pieces necessary to complete the Ln puzzle: the identification of novel transcriptional regulators and genes of unknown function, and the discovery and visualization of Ln storage in M. extorquens AM1. Growth phenotypes are reported for over 40 strains in methanol media containing and lacking Ln and reveal requirements for novel genes in both Ln-dependent and Ln-independent methanol growth. A detailed phenotypic characterization for strains lacking each component of the first 8 genes in the Ln transport cluster is described, including the ability of these strains to mutate or acclimate to allow Ln transport through a predicted secondary mechanism. Ln uptake was also quantified for several transport mutant strains. Finally, we show that M. extorquens AM1 not only incorporates Ln into the cytoplasm, but also stores Ln as crystalline cytoplasmic deposits. To our knowledge, this is the first demonstration of this behavior by a methylotrophic microorganism known to use Ln for metabolism.
RESULTS
XoxF produces formaldehyde in vivo
While XoxF can produce formate directly from methanol in vitro, XoxF contributes to formaldehyde production in vivo in M. extorquens AM1 (8). To expand these findings, strains lacking fae (encoding a formaldehyde-activating enzyme) and the different ADHs were constructed and tested for growth on solid La3+ medium containing both succinate and methanol as carbon sources (Fig. 1, Table 1). To confirm that loss of the Ca2+-dependent MxaFI-MeDH does not affect growth of the fae mutant when Ln are present, an fae mxaF double deletion mutant strain was tested as a control. Loss of xoxF1 alone allowed survival of the fae mutant strain, while additional loss of xoxF2 allowed growth similar to the wild-type strain (Table 1). Loss of neither exaF nor mxaF conferred methanol resistance to the fae mutant strain, indicating that MxaFI and ExaF are not major contributors to formaldehyde production in vivo when Ln are present.
Ability of ADH mutations to confer methanol resistance to an fae mutant strain on succinate and methanol solid medium with La3+.
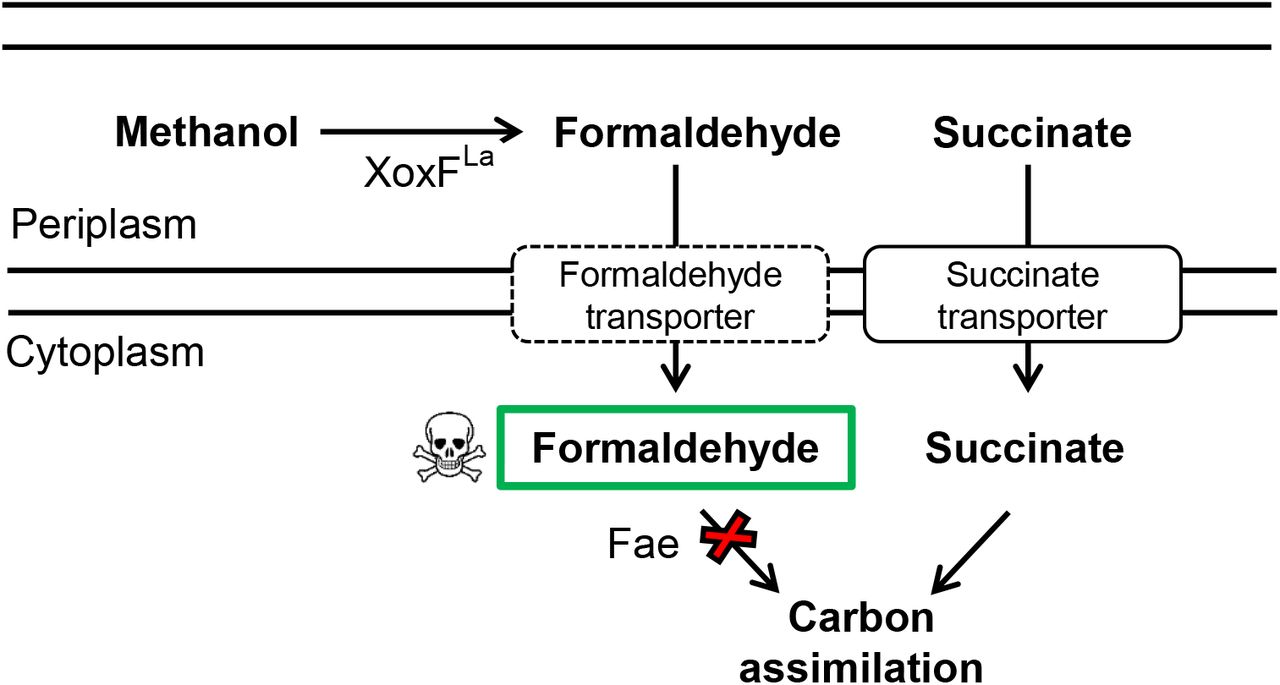
When coordinated to Ln, the XoxF methanol dehydrogenase is known to produce formaldehyde in vivo. When M. extorquens AM1 lacks fae as indicated by a red X, formaldehyde accumulates to toxic levels rendering strains unable to grow on solid media even in the presence of alternative substrates such as succinate. In the absence of mxaF, if a process required for XoxF function is disrupted by a transposon insertion, formaldehyde is reduced or eliminated, and growth can occur using succinate as a carbon and energy source. Dashed lines are used to represent the formaldehyde transporter as this function has not been demonstrated.
Identification of gene products that contribute to Ln-dependent methanol growth
A transposon mutagenesis study was designed to take advantage of the in vivo formaldehyde production capability of XoxF to identify genes required for Ln-dependent methanol oxidation. The strain used to conduct the mutant hunt contained a mutation in mxaF to make cells dependent on Ln for formaldehyde production, and a second mutation in fae, which would result in formaldehyde accumulation and death of the cells when methanol was provided as a substrate (Fig. 1). Transposon mutants with disruptions in genes involved in processes required for Ln-dependent methanol oxidation were selected for on medium containing succinate, methanol, La3+, tetracycline (Tc), and rifamycin (Rif). Transposon insertions that reduced or eliminated formaldehyde production allowed survival and colony formation since methanol resistant strains could use succinate for growth. In addition to genes required for methanol oxidation, this mutant hunt had the potential to isolate insertions in an unknown formaldehyde transport system, which would theoretically reduce formaldehyde levels in the cytoplasm (Fig. 1).
Over 600 transposon mutants were isolated, and their insertion locations mapped to the M. extorquens AM1 genome. A variety of colony sizes were evident indicating that in some mutants, formaldehyde production was eliminated while in others, it might only have been reduced. Since it is likely that a portion of these transposon mutants became methanol-resistant due to spontaneous second-site suppressor mutations and not a transposon insertion, only genes that were identified three or more times were considered for further analysis and are listed in Table 2. Among the genes identified were those with obvious roles in methanol oxidation such as PQQ biosynthesis and the xoxFGJ genes. Three different clusters predicted to function in cytochrome c biogenesis and heme export (cyc and ccm genes) (reviewed in (40)) were identified along with insertions in an mxaD homolog gene (41). Insertions were isolated in six of the Ln transport cluster genes including the ABC transporter components, the TonB-dependent receptor, and two putative exported proteins (Fig. 2A). Notably, insertions were not isolated in Ln-binding proteins, lanmodulin and META1_1781 (23, 24). Additional genes of unknown function were also discovered, including a LysR-type transcriptional regulator, a fused ABC transporter, a porin family protein, a putative homospermidine synthase, a cytosol aminopeptidase, and orf6 and orf7, which are located downstream of the pqqABCDE operon (42). Based upon the work detailed within, we propose to name the Ln transport cluster genes as lut for lanthanide utilization and transport (Fig. 2A).
Genes identified three or more times via transposon mutagenesis.
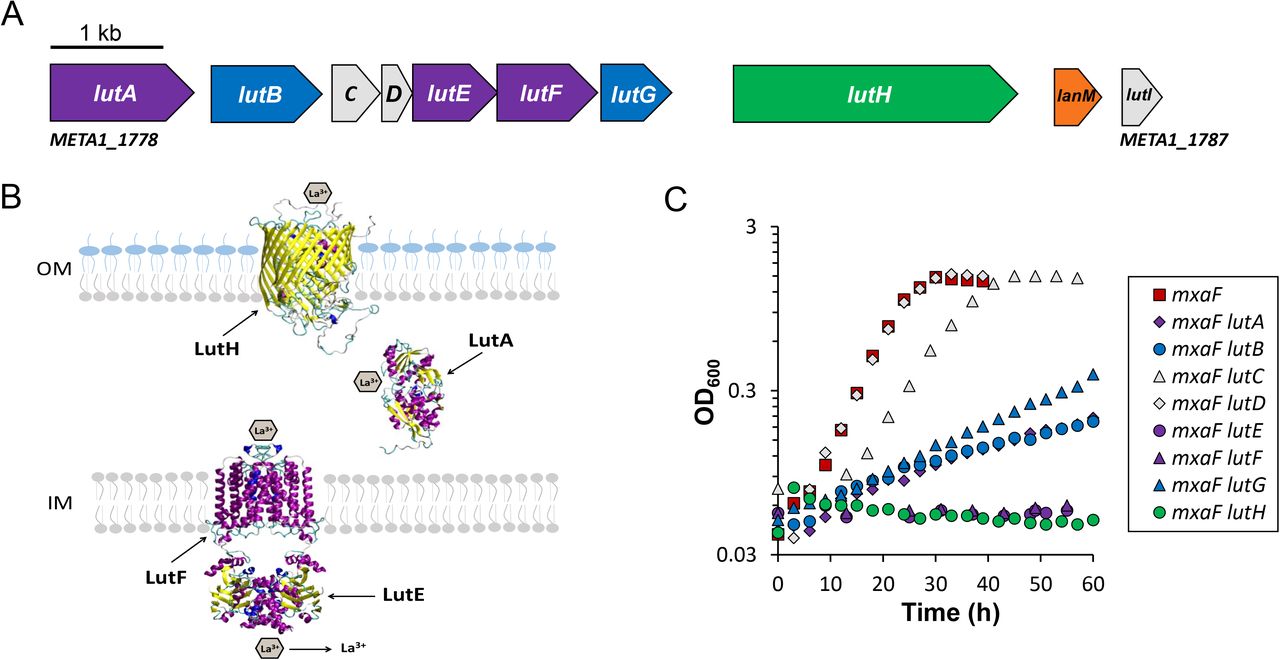
A) Genomic map of the genes contained in the Ln transport cluster (META1_1778 to META1_1787). Purple, genes encoding the ABC transport system; green, gene encoding the TonB-dependent transporter; blue, genes encoding putative exported proteins identified by transposon mutagenesis; gray, lanmodulin and additional exported genes in the Ln transport cluster not identified by transposon mutagenesis. B) Schematic representation of the proteins predicted to be involved in Ln transport. The three-dimensional structures of monomers of LutH and LutA were predicted using homology modeling (HHpredserver and MODELLER) (80) and homodimers of the ABC transporter were predicted using GalaxyHomomer (81). C) Growth of wild type and lut mutant strains grown in 125 mM methanol medium with 2 μM LaCl3. Graphs depict representative data from three biological replicates. Variation between replicates was < 5% except for mxaF lutC and mxaF lutD which had growth variances of 9 and 20% respectively as reported in Table 3.
Identification of novel components involved in methanol growth independent of Ln
To assess if the identified genes are specifically involved in Ln-dependent methanol oxidation or general methanol oxidation, 24 genes were chosen for reconstruction in mxaF and/or wild-type strain backgrounds. Methanol growth was assessed in the presence and absence of La3+ for all strains. As expected, deletion of genes in either of the two pqq biosynthesis operons (ΔpqqBCDE; ΔpqqF) eliminated Ln-dependent and independent methanol growth as did mutations in each of the three identified cytochrome c biogenesis and heme export clusters: cycK, heme lyase; ccmB, heme exporter; and ccmC, heme exporter (Table 3).
Growth parameters for strains grown in methanol medium with and without La3+.
Novel genes affecting methanol metabolism were identified (Table 3). First, a fused ABC-type transporter (META1_2359) mutant was unable to grow in methanol medium with or without La3+. After 85-100 h, second site suppressor mutations arose that allowed the strain to grow to similar culture densities as the wild-type strain but at a reduced growth rate. Second, META1_3908 encodes a putative leucyl aminopeptidase that shares 38% identity and 57% similarity with PepA from E. coli MG1655. Loss of META1_3908 resulted in a long growth lag (24 h without La3+; 15 h with La3+) and an approximate 60% decrease in growth rate in both conditions. The aminopeptidase appears to be in an operon based on overlap with META1_3909, a putative membrane protein of unknown function. Deletion of META1_3909 did not result in a growth defect in methanol media with or without La3+ (data not shown). Third, loss of hss (META1_2024) encoding a putative homospermidine synthase resulted in a relatively short lag and an approximate 35% reduction in growth rate in methanol medium containing or lacking La3+. Finally, while a LysR-type regulator (META1_0863) was hit nine times in the transposon mutagenesis study, loss of this LysR-type regulator only resulted in an approximate 10% decrease in growth rate in liquid methanol media. However, on solid methanol media, loss of META1_0863 had a more pronounced defect with colonies half the size of wild type (data not shown).
Identified genes specific to Ln-dependent methanol growth
It was previously shown that xoxF is required for growth in methanol medium that lacks Ln as XoxF is required for expression of the MxaFI-MeDH (14, 21). Deletions were constructed in the xoxG and xoxJ genes to confirm that xoxGJ are required for La3+-dependent, but not La3+-independent methanol growth in M. extorquens AM1 as recently shown for M. extorquens PA1 (22). In the absence of La3+, loss of xoxG resulted in a growth reduction of 21% when compared to wild-type growth, while the xoxJ growth defect was subtle (7% reduction) but with a 21 h lag (Table 3.) However, in methanol La3+ medium, loss of either xoxG or xoxJ was equivalent to loss of both xoxF1 and xoxF2 indicating that XoxGJ are essential for XoxF-dependent methanol oxidation as suggested by recent biochemical studies (43–45). Transcriptional reporter fusion studies to assess expression from the mxa promoter using a fluorescent reporter confirmed that unlike xoxF, neither xoxG nor xoxJ was required for expression of the mxa genes in succinate plus methanol medium (average relative fluorescence units (RFU)/OD600: wild type, 322.9 ± 62.5; xoxF1 xoxF2, 3.1 ± 1.3; xoxG, 313.9 ± 24.0; xoxJ, 512.5 ± 54.9) with the xoxJ mutant resulting in a 1.9-fold upregulation from the mxa promoter. These data are consistent with XoxF presence, but not XoxF activity, being required for expression of the mxa genes as reported for M. extorquens PA1 (22).
The MxaD homolog, META1_1771, shares 43% identity and 58% similarity to the M. extorquens AM1 MxaD protein, which has been suggested to facilitate interactions (directly or indirectly) between a MeDH and its cytochrome (41). Loss of the mxaD homolog did not cause a growth defect in methanol medium lacking La3+ but displayed an ~30% reduction in growth rate when La3+ was provided (Table 3). These data suggest an important but non-essential role for the MxaD homolog in Ln-dependent methanol oxidation.
Marker-less deletion strains lacking either orf6 or orf7 were constructed and methanol growth was assessed in the presence and absence of La3+. In methanol medium with La3+, growth of the orf6 mutant strain mirrored that of the xoxF1 xoxF2 double mutant while growth of the orf7 mutant was faster and like that of the xoxF1 single mutant (Table 3). Since these genes are downstream of the operon encoding the PQQ biosynthetic pathway (pqqABCDE), addition of PQQ was tested to see if it would rescue the growth defect seen for the orf6 and orf7 mutants. While 1 μM PQQ was able to rescue growth of the pqqBCDE and pqqF mutant strains, PQQ did not rescue growth of the orf6 or orf7 mutants (data not shown).
Finally, an outer membrane porin-like gene (META1_5071) was identified in the transposon mutant hunt. Insertions were constructed in wild-type and mxaF strain backgrounds and growth in the presence and absence of La3+ was assessed. A slight growth defect was observed in the wild-type background in methanol La3+ medium (~10% reduction). However, in the absence of mxaF when cells require Ln for growth, a further growth reduction was observed (~20% reduction) along with a growth lag of 21 h.
Genetic characterization of the lanthanide utilization and transport cluster
The increasing evidence that Ln-dependent enzymes are widely distributed among different microbial taxa and environments leads to the parallel question of how these elements are scavenged for use in living systems. Our transposon mutagenesis studies demonstrate that genes encoding homologs of the TonB- and ABC-dependent Fe3+ scavenging pathways play a role in methanol metabolism when Ln are present. In addition to the transport system homologs, two of the four putative exported proteins were identified as important for Ln-dependent growth. To facilitate a detailed genetic characterization of this gene cluster (Fig. 2A), mutations in individual genes from lutA through lutH were constructed in wild-type and mxaF mutant backgrounds and tested for growth in the presence and absence of La3+ (Table 3, Fig. 2C, Fig. S1). Mutants lacking individual transport cluster genes (lutABEFG) were complemented by expressing the respective gene in pCM62 (46) and growth similar to the wild-type strain was restored in each case (data not shown).
In the absence of mxaF, M. extorquens AM1 must obtain Ln for methanol growth (14) thus providing a condition to assess gene requirements for Ln utilization. Loss of the putative exported proteins that were not hit in the transposon mutant hunt either did not result in a growth defect (mxaF lutD) or decreased the growth rate by ~30% (mxaF lutC) (Table 3, Fig. 2C). Complementation of lutC was not tested so it is formally possible that the decrease in growth rate was due to polarity onto lutEFG. However, the lutC mutation was constructed such that adequate space for a rut site required for Rho-dependent polarity is not present (Table S1). In the mxaF strain background, loss of the remaining lut genes encoding putative exported or periplasmic components resulted in a significant growth defect. Specifically, loss of lutA (periplasmic binding component of ABC transporter) and the downstream putative exported protein encoded by lutB resulted in a ~88% reduction in growth rate, while strains lacking lutG displayed an ~81% reduction in growth rate (Fig. 2C, Table 3). The growth observed for these strains was not identified as second-site suppression or acclimation in our tests. These data show that while the periplasmic components of the Ln transport cluster are important to facilitate Ln utilization and transport, they are either not essential or have redundancy. This non-essentiality is in contrast with the requirement for the ATPase and membrane components of the ABC transporter (LutE and LutF, respectively), and the TonB-dependent receptor (LutH). Loss of lutE or lutF in the mxaF strain background did not allow growth (Fig. 2C). However, after ~200 h and 150 h respectively, second-site suppressor mutations arose which facilitated growth albeit 88% slower than that of the wild-type strain (Fig. S1, Table 4). The mxaF lutE and mxaF lutF strains retained the ability to grow in methanol with La3+ after succinate passage, indicating a genotypic change. In contrast, after 90-120 h, growth of the mxaF lutH strain occurred in only ~60% of the cultures (Fig. S1, Table 4). If growth did occur, this strain could grow without a lag once inoculated into fresh methanol La3+ medium and exhibited wild-type colony sizes on solid methanol medium similar to the wild-type strain (data not shown). However, if the culture was instead first streaked onto minimal succinate medium to obtain isolated colonies, then inoculated into or onto fresh methanol La3+ media (liquid or solid), the strain lost the ability to grow indicating that the original growth was due to acclimation, not suppressor mutations.
Growth parameters of suppressor and acclimation events.
As reported by Ochsner et al. (22), loss of lutH alone did not result in a growth defect in methanol La3+ medium consistent with the hypothesis that LutH is needed for transport of Ln into the cell and lack of Ln transport through the outer membrane enables growth with methanol via the Ca2+-MxaFI MeDH. For all other strains disrupted in the lut gene cluster, growth with a functional mxaF gene was similar to growth of strains lacking mxaF (Table 3). Intriguingly, loss of the membrane and ATPase components of the ABC transporter eliminated growth in the wild-type background (Fig. 2C, Table 4). However, suppressor mutations arose in the lutE and lutF mutant strains after 75-91 h. A second transposon mutant hunt revealed that insertions which restored the ability of the lutE mutant to grow on methanol La3+ medium mapped to the TonB-dependent receptor, lutH, consistent with expression of mxaF when cells are unable to sense Ln. These data suggest that Ln must enter the cytoplasm in order to upregulate expression of the xox and exaF ADHs and that Ln uptake into the periplasmic space may be enough to repress mxa expression. It is also possible that Ln must first enter the cytoplasm to be released from the predicted siderophore-like molecule (recently termed, lanthanophore (47)) to be incorporated into XoxF. One possibility does not exclude the other. These discoveries suggest new hypotheses to be tested regarding the Ln-switch.
Quantification of lanthanide uptake
The Arsenazo dye-based assay (48) was used to quantify La3+ levels in culture supernatants (spent media) from wild type; lutA, lutE, lutF (ABC transporter); and lutH (TonB-dependent receptor) mutant strains during growth (Fig, 3A). Strains were grown in methanol medium containing 2 µM La3+ and limiting succinate (3.75 mM) as lutA, lutE, and lutF mutant strains are unable to grow or grow poorly with methanol as a sole carbon source (Table 3). Under these conditions, no significant decreases in La3+ levels were observed for any strain during early-to mid-exponential phase (OD600 = 0.4) (Fig. 3B). As cultures continued to grow and succinate was likely depleted, significant differences in La3+ levels became apparent. At an OD600 of ~0.7, the supernatants of wild-type cultures contained 1.0 ± 0.1 µM La3+ (Fig. 3B). Similar levels were measured for the lutA, lutE, and lutF mutant strains (1.1 ± 0.1 µM La3+) suggesting Ln are still transported through the outer membrane. The lutA, lutE, lutF mutant strains were unable to grow to an OD higher than 0.68 (Fig. 3A), but after 8 h in stationary phase, the La3+ concentration in the supernatant continued to decrease to 0.7 ± 0.1 µM (data not shown). Levels of La3+ in the supernatants from lutH mutants grown to the same OD were significantly higher (1.5 ± 0.1 µM, Fig. 3B). Although a lutH mutant strain can grow to a similar density as the wild-type strain, it is hypothesized that unlike wild type, the lutH mutant grows using MxaFI for methanol consumption regardless of Ln presence, and therefore, differences in La3+ levels in the supernatant are expected. In stationary phase, La3+ levels in the supernatants from wild-type cultures were half of those found for the lutH mutant cultures (0.6 ± 0.1 µM and 1.3 ± 0.1 µM, respectively) (Fig. 3B). Decrease of La3+ in the lutH culture supernatants may reflect adsorption of La3+ onto the surface of the cells or interaction of La3+ with lipopolysaccharide, a phenomenon observed for other metals in different bacterial species (49). Additionally, the growth and acclimation data for the mxaF lutH strain suggests a secondary mechanism for Ln transport through the outer membrane (Table 3, Fig. S1).

A) Growth analysis of wild type (red circles), lutH encoding the TonB receptor (dark blue triangles), lutA encoding the periplasmic binding protein component of the ABC transporter (bright blue squares), lutE encoding the ATP binding component of the ABC transporter (light blue diamonds), and lutF encoding the transmembrane component of the ABC transporter (light blue circles), grown in the presence of limiting succinate (3.75 mM), methanol (125 mM), and 2 μM LaCl3. Data represent the average of three biological replicates with variances < 5%. B) La3+ concentration (μM) present in the supernatant at different optical densities (OD600) for wild type (red), lutH (dark blue), and lutA, lutE, lutF (gradient of light blue respectively). Each bar represents the average of three biological replicates with error bars showing the standard deviation. One-way analysis of variance (ANOVA) followed by a T-test was used to represent statistical significance. *The p-value is <0.005.
Visualization of La3+ accumulation in lut transporter mutants
To further investigate the roles for the components of the TonB-ABC transport system, location of La3+ was assessed by transmission electron microscopy (TEM) in the lutA, lutE, lutF, lutH mutant strains. TEM coupled with energy dispersive X-ray spectroscopy (EDS) has been used to determine the elemental composition of cellular inclusions (50, 51) while La3+ has been widely used as an intracellular and periplasmic stain for electron microscopy (52–54). Here, we show that La3+ can be directly identified by TEM if accumulated inside M. extorquens AM1 cells. Strains were grown in methanol medium containing 20 µM La3+ and limiting succinate (3.75 mM). Samples with post-fixation by OsO4 and staining with 2% uranyl acetate allowed the outer and inner membrane of the bacterial cells to be distinguished (Fig. 4A-E left subpanels), while visualization without fixation and staining enabled metal content analysis by removing the interaction between Os8+ and phosphate, which interferes with La3+ measurements (Fig. 4A-E right subpanels). Mutants lacking the TonB-dependent receptor (lutH) did not display La3+ deposits (Fig. 4A). In contrast, localized La3+ deposits were visualized in the periplasmic space in mutant strains lacking the ABC-transporter components (lutA, lutE, and lutF) as shown in Fig. 4B-E. EDS microanalyses confirmed these electron dense periplasmic deposition areas contained La3+ (Fig. 4F). Electron-dense deposits were also observed in the cytoplasm from wild-type cells grown with exogenous La3+ (Fig 4G). Taken together, these directly demonstrate the requirements for the TonB-dependent receptor and ABC transporter in Ln transport.

Thin sections of A) lutH, B) lutE C) lutF, D) lutA, and G) wild-type strains growth in minimal medium with 3.75 mM succinate, 125 mM methanol, and 20 µM LaCl3. White arrows indicate depositions of electron scattering material in the periplasm. Each panel depicts cells that were fixed with 2.5% glutaraldehyde, either post-fixed with OsO4 and stained with uranyl acetate to detect cell membranes (left subpanel), or without post-fixation and staining for elemental analysis (right subpanel). E) Magnification of the La-deposits localized in the periplasmic space from the lutA mutant strain; black arrows show the boundaries of the outer membrane and inner membrane. F) Energy-dispersive X-ray analysis of the electron-dense deposits observed inside the periplasm.
Expression of lutH is repressed by La3+
To test if expression of the of Ln uptake genes is regulated in a similar manner to Fe3+ uptake genes, a fluorescent transcriptional reporter fusion was used to monitor expression from the lutH promoter in methanol media. Addition of exogenous La3+ repressed expression 4-fold (RFU/OD600 = 210 ± 7 without La3+; RFU/OD600 = 47 ± 5 with La3+) suggesting that when Ln are in excess, transport is down-regulated. This is consistent with the mechanism of control for iron homeostasis (37, 39).
Ln are stored as cytoplasmic crystalline deposits
To determine if M. extorquens AM1 stores Ln, an mxaF mutant was grown in methanol medium with 10-times the standard concentration of La3+ (20 µM LaCl3) resulting in a growth rate of 0.15 ± 0.00 h−1. Six hours post stationary phase, cells were washed four times and sub-cultured into methanol medium lacking La3+ using Ln-depleted tubes (14). To ensure growth was not due to contamination, strains were streaked onto methanol solid medium without La3+. As shown in Fig. 5, a similar growth rate to that observed in the presence of La3+ was obtained (subculture #1, 0.16 ± 0.00 h−1) until the culture reached an OD600 of ~1.0. A slower growth rate was noted as growth continued to an OD of ~1.5. This culture was then washed and sub-cultured into fresh methanol medium lacking Ln as described above. Cells continued to grow but at a very impaired rate (0.01 ± 0.0 h−1) until they reached a peak OD of ~1.3. The use of Ln-depleted glassware reduces Ln levels such that growth of an mxaF mutant strain appears negligible for 100 h when grown in media lacking Ln (14). As these storage growth curves were carried out for >600 h, a control experiment was done to determine growth due to either background levels of non-MxaF ADH activity or due to leeching of residual Ln from the glassware. Growth of an mxaF mutant strain that had not been previously grown with Ln had slower growth than subculture #2 with a growth rate of 0.003 ± 0.000 h−1 (data not shown).

Optical density (OD600) measurements of cultures grown with excess LaCl3 (20 μM) (gray circles) until six hours post stationary phase. Cells were washed and reinoculated into La3+ free methanol medium in La3+ tubes (subculture #1, dark blue circles). After 6 hours in stationary phase, subculture #1 was washed and reinoculated into methanol medium without La3+ (subculture #2, light blue circles). Representative data from three biological replicates is shown. Growth variance was <5%.
To visualize Ln storage within the cell, cultures of the wild-type strain were grown in the presence and absence of 20 µM LaCl3, harvested at mid-exponential phase, and immediately fixed with glutaraldehyde. Samples without post-fixation and staining were analyzed for both visualization and metal content. Electron-dense deposits were observed in the cytoplasm from cells grown with exogenous La3+ (Fig. 6A-B). Samples were analyzed using EDS and corroborated that the dense deposits contained La3+ (Fig. 6C). When grown without La3+, only a few cells showed smaller electron dense areas (Fig. 6E-F); however, EDS analysis did not detect La3+ in these cases (Fig. 6G). These data demonstrate that La3+ can be stored in M. extorquens as metal deposits. Moreover, EDS analysis of electron dense areas from the wild-type strain grown with La3+ (Fig S2) determined a content of La (22.2 ± 1.0%), P (15.1 ± 2.1%), and oxygen (O) (51.1 ± 1.9%) suggesting Ln are complexed with phosphate. Traces of chloride (3.0%), calcium (2.2%), and aluminum (3.4%) ions were also detected. The copper, carbon, and silicon ion content from the support grids and embedding medium were not considered for metal content calculations. High-resolution transmission electron microscopy (HRTEM) images of the La deposits showed an atomic lattice (Moire fringes/pattern) indicating a crystalline nature (Fig. 6D). These results suggest that La is embedded in inorganic phosphate crystals which form the electron dense deposits observed in the cytoplasm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
TEM of ultrathin sections of wild-type cells grown with (A-B) and without (E-F) 20 µM La3+in minimal medium with 3.75 mM succinate and 125 mM methanol. C) and G) Elemental analysis of electron dense deposits was conducted using EDS on unstained ultrathin sections from cells grown with (C) and without (G) La3+. D) High-resolution transmission electron microscopy analysis of the wild-type strain revealing the presence of an atomic lattice in electron-dense areas that suggests La3+ is embedded in inorganic crystals inside the cell.
DISCUSSION
The Ln-dependent XoxF-MeDH has been shown to produce formaldehyde in vivo (8). Here we show that formaldehyde accumulation due to disruption of fae is lethal when tested on methanol and La3+ solid medium, which enabled a genetic selection to identify genes required for or involved in methanol oxidation by XoxF. Additionally, our phenotypic data are consistent with in vitro and in vivo work that suggests formate is the product of ExaF-mediated methanol oxidation (5, 8) and with our previous work that shows expression from the mxa promoter is repressed when Ln are present (8, 14).
The mutational analysis presented expands the identification of gene products contributing to methanol metabolism, Ln transport and utilization, and the characterization of genes with known or predicted roles in Ln metabolism. Using growth and transcriptional reporter fusion studies, we show that unlike xoxF, xoxG and xoxJ are not required for expression of the mxa genes but are likely required for XoxF activity as loss of either xoxG or xoxJ resulted in growth that mirrored the xoxF1 xoxF2 double mutant strain. Recent biochemical and structural analyses of XoxG suggest that this cytochrome is tuned specifically for light Ln, while XoxJ interacts with and may activate XoxF (44). In the methanotroph Methylomonas sp. strain LW13, loss of xoxG also results in a growth defect in methanol medium lacking Ln, suggesting an unknown role in metabolism in addition to functioning as a cytochrome for XoxF-mediated methanol oxidation (43). Our studies are consistent with an unknown role for XoxGJ in M. extorquens AM1 as strains lacking xoxG or xoxJ display a growth lag or reduction in growth rate. These data are in contrast to previous reports for M. extorquens PA1 and AM1, where loss of xoxGJ did not result in a growth phenotype in methanol medium lacking Ln (22, 42). Notably, the previous AM1 studies were carried out on agar plates where subtle growth defects may not be apparent and both published studies were conducted using a different growth medium.
Our transposon mutagenesis and growth studies lay the foundation for future work characterizing the roles for new methylotrophic gene products. A fused ABC-transporter (META1_2359) was found to be essential for methanol growth independent of Ln presence. Some possible functions for META1_2359 may include formaldehyde transport or PQQ export into the periplasm for incorporation into the ADHs. Leucyl aminopeptidases like META1_3908 are often involved in protein processing or turnover, yet some have been shown to exhibit DNA binding activity and facilitate regulation or site-specific recombination (55). Homospermidine synthases (hss) function in polyamine biosynthesis and their products (e.g. putrescine, spermidine) and have been implicated in roles as diverse as host pathogen interactions, biofilm formation, siderophore production, acid resistance, and free radical scavenging (56, 57). LysR-type regulators are members of a large family of transcriptional regulators that can be both activators and repressors of a wide variety of genes involved in diverse cellular processes (58). More work is required to identify specific roles for these genes in the metabolism of M. extorquens AM1.
Loss of the mxaD homolog, META1_1771, resulted in a growth defect only if La3+ was included in the medium. Based on its sequence similarity to MxaD, localization in the genome, and growth phenotypes, we propose to name META1_1771 as xoxD. Interestingly, upstream of xoxD is another mxaD homolog (META1_1772) and downstream is an mxaE homolog (META1_1770). These genes were not identified in the transposon mutant hunt and were not characterized in this study.
Before the existence of Ln-dependent MeDHs were known, mutations were constructed in orf6 and orf7 due to their proximity to C1 genes and it was concluded that these genes likely did not have a role in C1-metabolism as they grew on methanol medium in the absence of Ln (42). Results here clearly suggest that orf6 and orf7 are important for Ln-dependent methylotrophy and more work is needed to determine their specific roles.
Characterization of the lut operon is consistent with the observations recently reported for M. extorquens PA1 (22) where a TonB-ABC transport system contributes to Ln transport into the cytoplasm. It has been proposed that Ln may be acquired via lanthanophores (47), which enter through a TonB-dependent receptor (22, 23). It may be that secondary mechanisms exist to take up Ln chelated by phosphates or citrate, compounds which are present in Methylobacterium PIPES (MP) medium (59). It is possible that these complexes pass through the outer membrane via a porin, though that is speculation at this point. Of interest, META1_5017 is a porin family protein that was identified in our transposon mutagenesis studies, and loss of this gene resulted in a 21 h lag and an approximate 20% reduction in growth rate only in the mxaF mutant background. This phenotype would be consistent with a reduction in Ln import. It may be that in the lutH mutant, non-siderophore bound Ln slowly enter through META1_5017 porin and once a threshold is reached, trigger expression and function of the Ln-dependent ADHs. This may explain the acclimation of the mxaF lutH strain after ~100 h but more work is needed to confirm this hypothesis. Since transposon mutants disrupted in the genes encoding lanmodulin or the Ln-binding protein downstream of lanmodulin were not isolated, our observations are consistent with the findings in M. extorquens PA1 that suggest a non-essential or redundant role for these genes in Ln metabolism (22). Notably, the mxaF fae transposon mutagenesis study did not identify obvious gene candidates for lanthanophore biosynthesis though over 600 insertions were mapped to the genome. This may indicate that more than one lanthanophore is produced by the cell or that the lanthanophore has an essential role that is not yet understood. Additionally, we show that like the paradigm for regulation of siderophore mediated Fe3+ uptake, expression from the TonB-dependent receptor promoter, lutH, is repressed by Ln.
Our transport, TEM, and EDS analyses are consistent with LutH facilitating Ln transport into the periplasm and the ABC transport system facilitating Ln transport into the cytoplasm. La3+ concentrations found in the supernatant from strain variants lacking transport system components suggest that once in the periplasm, significant concentrations of La3+ do not go back outside of the cell as ABC transporter mutants showed uptake of La3+ from the medium similar to the wild-type strain. Intriguingly, TEM and EDS studies with ABC transporter mutant strains, demonstrated localized accumulation of Ln in the periplasmic space. It is not yet clear how or why Ln accumulate in specific areas rather than appear diffused throughout the periplasm. Taken together, our phenotypic growth, transport, and visualization studies suggest that Ln must enter the cytoplasm for the xox and exaF ADHs to be expressed and that Ln uptake into the periplasmic space may be enough to repress mxa expression. This work will facilitate new exploration of the Ln-switch.
It has been observed that bacteria such as Bacillus licheniformis (60), Myxococcus xanthus (54), and Pseudomonas aeruginosa (61) can effectively adsorb La3+ in mineral form onto their cell surface when Ln are at high concentrations (mM). Here, TEM and EDS analyses demonstrate that M. extorquens AM1 stores Ln in the cytoplasm in crystal form. This is consistent with recent studies for the non-methylotroph Thermus scotoductus SA-01 which showed that Eu3+ can accumulate in the cytoplasm. However, these studies were conducted using very high Eu3+ concentrations that are not typically found in nature (51). Further, a role for Eu3+in the metabolism of T. scotoductus SA-01 remains to be defined. For many bacteria, biomineralization is a mechanism used to cope with toxicity of different metals, manage waste products, sense and change orientations in accordance with geomagnetic fields, and store important cations for growth (62–67). It has been reported that some microorganisms store polyphosphate as volutin or metachromatic granules, and these granules are often complexed with cations like Mg2+ and Ca2+ (68–71). It is not yet known if M. extorquens AM1 stores Ln complexed to polyphosphate, however, the ratios of P, O, and La detected in our studies are consistent with Ln phosphates. Detailed studies are necessary to define the exact chemical structure of Ln storage deposits in M. extorquens AM1. Our current findings bring exciting implications for bacterial metabolism and cell biology, and for the development of bioremediation and biometallurgy strategies for Ln recovery.
MATERIALS AND METHODS
Bacterial strains and cultivation
Strains and plasmids used in this study are listed in Table S2. E. coli strains were cultivated in Lysogeny Broth (LB) medium (72) (BD, Franklin Lakes, NJ) at 37°C. M. extorquens AM1 strains were grown in Methylobacterium PIPES [piperazine-N,N’-bis(2-ethanesulfonic acid)] (MP) media (59) supplemented with succinate (15 mM) and/or methanol (125 mM) as described (14) unless otherwise stated. Conjugations took place on Difco Nutrient Agar (Thermo Fisher Scientific, Waltham, MA). Cultures were grown in round-bottom polypropylene or borosilicate glass culture tubes, or 250 mL polypropylene Erlenmeyer flasks (Thermo Fisher Scientific, Waltham, MA). If glass tubes or flasks were used to culture bacteria, they were pretreated to remove Ln as previously described (14). Liquid cultures were grown at 29°C and shaken at 200 and 180 rpm in Innova 2300 and Excella E25 shaking incubators (Eppendorf, Hamburg, Germany), respectively. LaCl3 was supplemented to a final concentration of 2 or 20 μM when indicated. To prepare PQQ, methoxatin disodium salt (Santa Cruz Biotechnology, Dallas, Tx) was dissolved in deionized water at pH 12-13 and filter sterilized. When necessary, antibiotics were added at the following concentrations: rifamycin (Rif, 50 µg/mL), tetracycline (Tc, 10 µg/mL for LB, 5 µg/mL for MP or 10 µg/mL when used together with Rif), kanamycin (Km, 50 µg/mL), ampicillin (Ap, 50 µg/mL).
Plasmid and strain construction
Primers used for plasmid and strain construction are listed in Table S1. The allelic exchange plasmid pHV2 was constructed by cloning the sacB gene from pCM433 (73) into the PscI site of pCM184 (74) in the same orientation as the Tc resistant gene. Insertion and orientation of sacB was confirmed by colony PCR. The lutH transcriptional reporter fusion was constructed by cloning the promoter region of lutH into the AclI and EcoRI sites upstream of a promoter-less venus gene in pAP5 (21). To create overexpression constructs for complementation studies, individual genes in the lut operon (lutA, lutB, lutE, lutF, and lutG) were cloned into the KpnI and SacI sites downstream of a Plac promoter in pCM62 (46). Diagnostic PCR was used to confirm successful integration of inserts. Plasmids were maintained in E. coli TOP10 (Invitrogen, Carlsbad, CA).
Gene deletions were constructed using pCM184 or pHV2 as previously described (14) except 5% sucrose was added for counter selection against single crossovers (73) when using pHV2. Plasmids were conjugated into M. extorquens AM1 via biparental mating using E. coli S17-1 (75) or triparental mating using E. coli TOP10 (Invitrogen, Carlsbad, CA) and E. coli harboring the conjugative plasmid pRK2013 as described (14). When indicated, the Km resistance cassette was resolved using pCM157 to achieve marker-less deletions (74).
Transposon mutagenesis
Suicide vector pCM639 carrying a mini transposon ISphoA/hah-Tc (76) was conjugated into mxaF fae and lutE strain backgrounds via triparental mating as described (14, 77). Dilutions of the mating mixtures were plated onto MP succinate (15 mM) plus methanol (50 mM) La3+ medium for the mxaF fae strain background and MP methanol (125 mM) La3+ medium for the lutE strain background. Media contained 10 µg/mL Tc to select for successful integration of the mini transposon into M. extorquens AM1 chromosome and 50 µg/mL Rif to counter select against E. coli strains bearing pCM639 or pRK2013. Plates were incubated for 5-7 days at 29°C. Transposon mutant colonies were streaked onto MP succinate plus methanol La3+ Tc (mxaF fae) or MP methanol La3+ (lutE) medium for downstream studies.
Location of transposon insertions
To identify the transposon insertion sites, genomic DNA was isolated using Qiagen’s DNeasy UltraClean microbial kit (Qiagen, Germantown, MD). Degenerate nested PCR was performed as described (77, 78) with the following exceptions: PCR reactions contained 1 µM of each primer, 0.05 U/µL Dream Taq (Thermo Fisher Scientific, Waltham, MA), and 5% dimethyl sulfoxide. Modifications to the PCR amplification parameters included 2 minutes for the initial denaturation at 95°C, 6 cycles of annealing at 40°C followed by 25 cycles of annealing at 65°C for the first PCR reaction, and 30 cycles of annealing at 65°C for the second PCR reaction. PCR products were purified using a Qiagen PCR purification kit (Germantown, MD). Sequence analysis was performed using TransMapper, a Python-based software developed in-house to identify transposon insertion locations and map them to the M. extorquens AM1 genome. Insertion locations were visualized using the SnapGene Viewer (GSL Biotech LLC, Chicago, IL).
Phenotypic analyses
Growth phenotypes were determined on solid or in liquid MP media using a minimum of three biological replicates. On solid media, colony size was scored after four days. Growth curve analysis was conducted at 29°C in an Excella E25 shaking incubator (New Brunswick Scientific, Edison, NJ) using a custom-built angled tube rack holder as previously described (14). Optical density (OD600) was measured at 600 nm using a Spectronic 20D spectrophotometer (Milton Roy Company, Warminster, PA). For strains with extended growth lags, suppression and acclimation was assessed. Strains from the growth curves were streaked onto methanol La3+ medium after they reached stationary phase. If the parent stock strain did not grow on methanol La3+ and the strain post-growth curve grew, acclimation versus suppression was tested. Strains were passaged from the methanol medium plate to a succinate medium plate. After colonies grew on succinate medium, they were streaked back onto methanol La3+ medium. If strains retained the ability to grow on methanol medium, it was concluded that growth was due to a suppressor mutation. If strains lost the ability to grow on methanol medium after succinate passage, it was concluded growth was due to acclimation and not a genetic change.
For Ln storage growth experiments, mxaF deletion strains were grown with 20 µM LaCl3 until six hours after entrance into stationary phase. Cells were centrifuged and washed four times with MP medium lacking La3+ and a carbon source, and sub-cultured into fresh MP methanol medium without La3+. Six hours post stationary phase, the cultures were streaked onto MP methanol medium with and without La3+ to check for contamination. Cultures were again washed and sub-cultured as described above. This process was repeated for two rounds of growth without La3+.
Transcriptional reporter fusion assays
M. extorquens AM1 strains carrying mxa and lutH transcriptional reporter fusions (Table S2) which use venus (79) as a fluorescent reporter were grown in MP medium supplemented with methanol only or methanol and succinate with and without La3+ as indicated in the text. Expression was measured as relative fluorescent units (RFU) using a SpectraMax M2 plate reader (Molecular Devices, Sunnyvale, CA) and normalized to OD600 as previously described (14).
La3+ depletion during M. extorquens AM1 growth
Overnight cultures of wild type, lutA, lutE, lutF, and lutH mutant strains were inoculated 1:50 into 250 mL polycarbonate flasks (Corning Inc., Corning, NY) containing 75 mL of MP medium (59). Succinate (3.75 mM) and methanol (125 mM) were added as carbon sources with 2 μM LaCl3. Flasks were incubated at 28°C at 200 rpm in Innova 2300 shaking incubators (Eppendorf, Hauppauge, NY) for 44 h. To monitor La3+ depletion during M. extorquens AM1 cultivation, the Arsenazo III assay was used as previously described (48). 5 mL samples were collected at 4 different time points (OD600 of 0.04, 0.4, 0.7, and 1.5). The concentration of La3+ remaining in the supernatant was calculated using the calibration curve prepared as previously described (48). A control of 3 uninoculated flasks containing MP medium with 2 μM LaCl3 were considered to determine La3+ adsorption by the flasks which was subtracted from the culture measurements. The initial concentration of La3+ in the media (before growth) was measured using the Arsenazo III assay in the same way as described above. Significant differences between depletion of La3+ by different strains were calculated using One-way ANOVA followed by a T-test.
Cellular locations of Ln visualized using Transmission Electron Microscopy (TEM)
Sample preparation for TEM: wild-type, lutA, lutE, lutF, and lutH mutant strains were grown in MP medium containing 125 mM methanol and 3.75 mM succinate as carbon sources with or without the addition of 20 μM LaCl3 until they reached an OD of ~0.6. 3 mL of cells were harvested by centrifugation for 3 min at 1500 × g at room temperature and fixed for 30 min in 1 mL of 2.5% (v/v) glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA) in 0.1 M cacodylate buffer (Electron Microscopy Sciences, Hatfield, PA) at room temperature. After fixation, cells were pelleted by centrifugation for 3 min at 1500 × g at room temperature and washed with 1 mL of 0.1 M cacodylate buffer. Cell pellets were embedded in 2% (w/v) agarose and washed three times with 0.1 M cacodylate buffer. When indicated, pellets in agarose blocks were post-fixed for 30 min in 1% osmium tetroxide in 0.1 M cacodylate buffer. Samples were washed three times with 0.1 M cacodylate buffer, dehydrated in acetone, and embedded in Spurr resin (Electron Microscopy Sciences, Hatfield, PA). Blocks were polymerized at 60ºC for 48 h. 70 nm sections were obtained with a Power Tome XL ultramicrotome (RMC Boeckeler Instruments, Tucson AZ), deposited on 200 mesh carbon coated grids and stained with 2% uranyl acetate (Electron Microscopy Sciences, Hatfield, PA). To assess the presence of La3+ by EDS, sections were left unstained. To image the distribution of cellular La, a TEM JOEL 1400 Flash (Japan Electron Optics Laboratory, Japan) was used. Detection of La in the cells and high-resolution imaging were done with a JEOL 2200FS (Japan Electron Optics Laboratory, Japan) operated at 200kV. X-ray energy dispersive spectroscopy was performed using an Oxford Instruments INCA system (Abingdon, United Kingdom).
FUNDING SOURCES
This material is based upon work supported by the National Science Foundation under Grant No. 1750003 and by a California State University Program for Education and Research (CSUPERB) Joint Venture Grant. P.R-J. was supported by the National Science Foundation under Grant No. 1750003. EA and F.Y. were supported by the National Science Foundation Research Initiative for Scientific Enhancement (RISE) award under Grant No. R25GM071381. F.Y. was also supported by the National Institute of Health Maximizing Access to Research Careers Undergraduate Student Training in Academic Research (MARC U-STAR) award under Grant No. 4T34GM008253. Funding for transposon DNA isolation, PCR, and sequencing was provided by San José State University through the Department of Biological Sciences.
ACKNOWLEDGMENTS
We would like to thank Dr. Lena Daumann and Dr. Nathan Good for critical review of this manuscript. We would like to thank San José State University General Microbiology students who isolated transposon mutants and purified genomic DNA for sequencing as part of a class research project. The mutant hunt in this study was inspired by Dr. Elizabeth Skovran’s undergraduate research mentor, Dr. Marc Rott at the University of Wisconsin-LaCrosse who used transposon mutagenesis in the teaching lab to identify genes involved in butanol oxidation in Rhodobacter sphaeroides. We would like to thank Tim Andriese for assistance with sequencing of the transposon mutant DNA and all Skovran lab members for assistance with growth curves and transcriptional reporter fusion assays. TEM work was done at the Center for Advanced Microscopy, Michigan State University. We would like to thank Dr. Alicia Withrow for invaluable assistance with TEM experiments and Dr. Xudong Fan for his expertise using TEM-EDS. E.S., N.C.M-G., P.R-J., and H.N.V. directed experiments. E.S., N.C.M-G., P.R-J., and H.N.V. wrote the manuscript. P.R-J. conducted microscopy and Ln transport experiments. H.N.V, G.A.S, J.C., and E.C carried out transposon mutagenesis studies. H.N.V., G.A.S., R.C., J.C., C.R., E.M.A., E.C., N.F.L., and F.Y. constructed strains and conducted growth experiments. R.C., C.H., and G.A.S. conducted expression studies. R.T.N created TransMapper.
REFERENCES
- 1.↵
- 2.
- 3.↵
- 4.↵
- 5.↵
- 6.
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.
- 17.↵
- 18.
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.
- 29.↵
- 30.↵
- 31.
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.
- 64.
- 65.
- 66.
- 67.↵
- 68.↵
- 69.
- 70.
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵




