

SUMMARY
Lipid rafts form signaling platforms on biological membranes with incompletely characterized role in immune response to infection. Here we report that lipid raft microdomains are essential components of the phagolysosomal membrane of macrophages. Genetic deletion of the lipidraft chaperons flotillin-1 and flotillin-2 demonstrate that the assembly of both major defense complexes vATPase and NADPH oxidase on the phagolysosomal membrane requires lipid rafts. Furthermore, we discovered a new virulence mechanism leading to the dysregulation of lipid-raft formation by melanized wild-type conidia of the important human-pathogenic fungus Aspergillus fumigatus. This results in reduced phagolysosomal acidification. Phagolysosomes with ingested melanized conidia contain a reduced amount of free Ca2+ ions as compared to phagolysosomes with melanin-free conidia. In agreement with a role of Ca2+ for generation of functional lipid rafts, we show that Ca2+-dependent calmodulin activity is required for lipid-raft formation on the phagolysosome. We identified a single nucleotide polymorphism in the human FLOT1 gene that results in heightened susceptibility for invasive aspergillosis in hematopoietic stem-cell transplant recipients. Collectively, flotillin-dependent lipid rafts on the phagolysosomal membrane play an essential role in protective antifungal immunity in humans.
INTRODUCTION
Many bacteria manipulate professional phagocytes to counteract immune responses. For example, Mycobacterium spp., Legionella spp., and Listeria spp., withstand intracellular degradation after ingestion by phagocytes (Cambier, Falkow et al., 2014, Carvalho, Sousa et al., 2014, Ensminger, 2016). This is accomplished by their ability to interfere with the formation of a functional hostile phagolysosome (PL). Pathogenic fungi have also developed mechanisms to avoid being killed after they are taken up by phagocytes. The encapsulated yeast, Cryptococcus neoformans, damages phagosomal membranes, deploys potent antioxidant mechanisms and manipulates phagosomal maturation, which enables the fungus to survive and replicate inside phagosomes (Levitz, Nong et al., 1999, Smith, Dixon et al., 2015, Tucker & Casadevall, 2002, Zaragoza, Chrisman et al., 2008). Furthermore, C. neoformans can induce non-lytic exocytosis, in which the ingested pathogen is expelled from the phagocyte without lysing it (Alvarez & Casadevall, 2006, Ma, Croudace et al., 2006, Nicola, Robertson et al., 2011). Other fungi, such as the yeast forms of Candida spp. and Histoplasma capsulatum employ mechanisms to establish an intracellular lifecycle or to avoid killing inside the PL, respectively (Fernandez-Arenas, Bleck et al., 2009, Newman, Gootee et al., 2006, Seider, Brunke et al., 2011, Strasser, Newman et al., 1999). For example, H. capsulatum yeast cells reduce phagosomal acidification by releasing the saposin-like protein CBP that reduces vacuolar ATPase (vATPase) accumulation on the phagosomal membrane (Batanghari, Deepe et al., 1998, Woods, 2003). Rhizopus spp. establish intracellular persistence inside alveolar macrophages. Lack of intracellular swelling of Rhizopus conidia results in surface retention of melanin, which induces phagosome maturation arrest through inhibition of LC3-associated phagocytosis (LAP) (Andrianaki, Kyrmizi et al., 2018).
In line, we previously reported that dihydroxynaphthalene (DHN) melanin of conidia (spores) of the important pathogenic fungus Aspergillus fumigatus that is predicted to cause more than 200.000 life-threatening infections worldwide (Brown, Denning et al., 2012, Kosmidis & Denning, 2015), is responsible for establishing a non-hostile intracellular niche inside phagocytes by inhibiting phagolysosomal acidification, and as a consequence killing of conidia, LAP and apoptosis of phagocytes (Akoumianaki, Kyrmizi et al., 2016, Heinekamp, Thywissen et al., 2012, Schmidt, Vlaic et al., 2018, Thywissen, Heinekamp et al., 2011, Volling, Thywissen et al., 2011). The grey-green surface pigment consisting of dihydroxynaphthalene (DHN) melanin (Jahn, Koch et al., 1997, Langfelder, Streibel et al., 2003, Tsai, Chang et al., 1998) plays a pivotal role in this process because melanin ghosts, i.e., isolated pigment particles, led to reduced acidification to the same extent as wild-type conidia and impaired LAP, whereas conidia from the pigmentless pksP mutant showed full acidification of PLs and full activation of LAP (Akoumianaki et al., 2016, Thywissen et al., 2011). Furthermore, the absence of conidial melanin in the pksP mutant results in a higher phagocytosis rate, faster endocytotic processing, and increased production of pro-inflammatory cytokines and chemokines by phagocytes most likely due to better recognition of pksP conidia by the dectin-1 receptor (Chai, Netea et al., 2010, Luther, Torosantucci et al., 2007, Mech, Thywissen et al., 2011, Thywissen et al., 2011). Further data indicated that DHN-melanin is able to abrogate the release of free Ca2+ ions to the peri-phagosomal area thereby inhibiting calmodulin activity and impairing phagosome function (Kyrmizi, Ferreira et al., 2018). However, the downstream effector pathways regulating phagosome biogenesis and pathogen killing are incompletely characterized.
For fungal pathogens, the molecular mechanisms involved in the dysregulation of PLs and, furthermore, many aspects of phagosome maturation are unsolved. Here, we add novel insight into both aspects by the finding of lipid-raft microdomains as being essential for phagosome maturation, defense against invasive aspergillosis and also as a target of pathogens. We discovered an unprecedented virulence mechanism depending on the inhibition of flotillin-dependent lipid raft formation in phagolysosomal membranes by conidial melanin.
RESULTS
Reduced phagolysosomal acidification increases conidia-induced damage of macrophages
If a macrophage ingests an A. fumigatus conidium and fails to kill it, the conidium swells and germinates to produce a hypha that then pierces the macrophage and eventually kills it. We used the vATPase inhibitor, bafilomycin A1, to determine whether inhibition of phagolysosomal acidification affects the extent of macrophage damage. Wild-type conidia caused considerable macrophage damage whereas pksP mutant conidia lacking DHN-melanin caused significantly lower host cell damage (Fig. 1A). Abolishing phagolysosomal acidification with bafilomycin A1 increased the macrophage damage caused by both wild-type and pigmentless pksP conidia. Importantly, in the presence of bafilomycin A1, wild-type and pksP conidia caused virtually identical host cell damage. These results indicate that the capacity of wild-type A. fumigatus conidia to reside in a PL with reduced acidification enhances the capacity of the organism to survive within the phagocyte and eventually to kill it.

A) Cell damage monitored by 51Cr release from RAW264.7 macrophages infected with A. fumigatus wild-type or pksP conidia; data are represented as mean ± SD; p < 0.05. B) RAW264.7 macrophage cell had simultaneously phagocytosed melanized wild-type conidia (labeled with FITC, green) and non-pigmented pksP conidia (labeled with CFW, blue). Acidified phagolysosome stained with LysoTracker Red. Scale bar = 5 μm C) RAW264.7 macrophage had simultaneously phagocytosed a wild-type conidium (FITC-labeled) and a pksP conidium (CFW-labeled). Localization of cytoplasmic vATPase subunit V1 to the phagolysosomal membrane was monitored by immunofluorescence. Scale bar = 5 μm D) Western blot analysis for detection of vATPase V1 subunit using an ATP6V1B antibody. Cytoplasmic and membrane fractions of macrophages infected with wild-type or pksP conidia were analyzed. As controls, the cytoplasm and membrane specific proteins GAPDH and COXIV, respectively, were detected. E) Western blot data (from D) were quantified densitometrically to determine the ratio of cytoplasmic to total ATP6V1B subunit content of RAW264.7 macrophages infected with wildtype or pksP conidia; data show mean values ± SD; p < 0.05.
Reduced acidification is specific for PLs containing melanized conidia and correlates with reduced assembly of functional vATPase in the phagolysosomal membrane
To investigate whether the inhibition of phagolysosomal acidification by wild-type conidia is a global phenomenon or is limited to PLs that contain melanized conidia, macrophages were loaded with LysoTracker Red, simultaneously infected with melanized wild-type and pigmentless pksP conidia. Acidified PLs were identified by their red fluorescence. We found that when wildtype and pksP conidia were ingested by the same macrophage, there was no acidification of the PL containing a wild-type conidium, but clear acidification of the PL containing the pksP conidium (Fig. 1B).
We hypothesized that wild-type conidia-containing phagolysosomal membranes contain less active vATPase. Active vATPase consists of two complexes; the V0 complex is integrated into the membrane and, when assembled with the cytoplasmic V1 complex, shuttles protons across the lipid bilayer (Cotter, Stransky et al., 2015). To analyze the binding of cytoplasmic V1 to the V0 complex the localization of the V1 complex to the membranes of PLs that contained either wildtype or pksP conidia was monitored by immunofluorescence. In the same macrophage, we only found faint staining of the phagolysosomal membrane that surrounded wild-type conidia (Fig. 1C). By contrast, there was a strong signal for V1 at the phagolysosomal membrane surrounding pksP conidia. To further quantify the amount of assembled vATPase, immunoblotting was used to determine the relative amount of the V1 complex in the membrane and cytosolic fractions of macrophages containing wild-type conidia and pksP conidia to calculate the ratio of cytosolic V1 to the total amount of V1 subunits. There was more of the V1 complex in the cytosolic fraction in macrophages infected with wild-type conidia relative to macrophages infected with pksP conidia (Fig. 1D, E). These results indicate that wild-type conidia-containing PLs contain less assembled vATPase complex and thus reduced acidification. This mechanism is specific to the PL in which melanized conidia are located.
Melanized conidia interfere with formation of lipid-raft microdomains in PLs
To examine whether the formation of lipid-raft microdomains, which are rich in cholesterol and sphingolipids (Lingwood & Simons, 2010), in the PL membrane was disturbed when DHN-melanin-containing conidia were ingested, the presence of several lipid-raft marker lipids was analyzed. For this purpose, macrophages were infected with melanized and pksP conidia and then stained with fluorescently labeled cholera toxin subunit B (CTB) to label the sphingolipid GM1 ganglioside, whose accumulation at distinct membrane sites is characteristic of lipid rafts and which was used as an initial indicator for lipid-raft microdomains (Ledeen & Wu, 2015). Interestingly, in both MH-S and RAW264.7 macrophages that had ingested melanized conidia, there was only minimal CTB staining of the phagolysosomal membrane, despite the strong CTB staining of the macrophage cytoplasmic membrane (Fig. 2A, B). By contrast, in macrophages that had ingested pksP conidia, distinct CTB staining of both the phagolysosomal and cytoplasmic membranes was visible (Fig. 2A, B).

A) Murine RAW264.7 macrophages stained for GM1 ganglioside (CTB-Alexa 647, red) were infected with FITC-labeled wild-type or pksP conidia. B) Quantification of GM1 positive phagolysosomes (PLs) in MH-S and RAW264.7 macrophages after infection with wild-type or pksP conidia. Data are represented as mean ± SD; p < 0.05. C) Cholesterol in conidia-containing PLs of RAW264.7 macrophages was detected by filipin III staining. Conidia were stained with FITC; pksP conidia-containing PLs display a distinct ring-like structure of cholesterol (indicated by white arrows), which was lacking in wild-type conidia-containing PLs. See also Figures S1, S2, S3 and S4 and supplemental videos 1 and 2. D) Ca2+ ions in PLs containing wild-type and pksP conidia stained with Fluo-4 AM. E) Acidification of PLs and GM1 staining of PLs containing the indicated conidia upon addition of calmodulin inhibitor W7.
To further substantiate this finding, the cholesterol-specific fluorescent probe filipin III and the lipophilic dye DiD were used. As shown by confocal laser scanning microscopy (CLSM), both filipin III (Fig. 2C) and DiD (Fig. S1) weakly labeled membranes surrounding melanized conidia while strongly labeling membranes enclosing pksP conidia. These results confirm that the presence of DHN-melanin on the ingested conidium affects the lipid composition of the phagolysosomal membrane.
To determine the relationship between lipid rafts and phagolysosomal acidification, we demonstrated that PLs containing melanized conidia were neither acidified nor contained GM1-stained lipid-raft microdomains, whereas PLs containing pksP conidia contained acidified endosomal and lysosomal vesicles that strongly colocalized with lipid-raft microdomains (Fig. S2). The accumulation of lipid-raft microdomains in phagolysosomal membranes enclosing pksP conidia was also visualized as 3D reconstruction (supplemental Video S1). By contrast, such lipid-raft spots were less prominent in phagolysosomal membranes enclosing melanized wildtype conidia (supplemental Video S2).
In additional experiments, we found that melanin ghosts had the same capacity as wild-type conidia to reside in non-acidified PLs with reduced formation of lipid-raft microdomains (Fig. S3). Collectively, these data indicate that lipid-raft microdomain formation correlates with the acidification of the PL and that these processes are reduced in PLs containing melanized conidia or melanin ghosts.
To determine whether reduced lipid raft formation influences PL acidification, macrophages were treated with methyl-β-cyclodextrin (MβCD), which depletes the cells of cholesterol and consequently blocks the formation of lipid rafts (Keller & Simons, 1998). MβCD treatment reduced the total cellular cholesterol content to less than 20 % compared to the untreated control (Fig. S4). Exposure to MβCD resulted in a complete absence of acidification of the PL as well as almost all of the endosomal and lysosomal vesicles (Fig. S4). Although approximately 10 % of PLs containing melanized and pksP conidia still stained positive for GM1, acidification of these PLs was almost completely abolished (Fig. S4). Thus, the presence of lipid-raft microdomains in the phagolysosomal membrane contributes to proper acidification.
Inhibition of calmodulin activity reduces microdomain formation in the phagolysosomal membrane
Previously we showed that melanized conidia reduce the release of free Ca2+ ions from the phagosome lumen to the peri-phagosomal area, thereby preventing activation of calmodulin and LAP (Kyrmizi et al., 2018). Thus, it was conceivable that Ca2+ sequestering by DHN-melanin also dysregulates lipid-raft microdomain formation. To test this assumption we first tested the presence of Ca2+ ions in PLs containing melanized and pksP conidia using Fluo-4 AM. Fluorescence microscopy indicated a much higher content of Ca2+ ions in pksP-containing PLs compared with wild-type conidia-containing PLs (Fig. 2D). Next, we inhibited calmodulin activity by addition of the inhibitor W7 that led to reduced formation of lipid-raft microdomains and reduced acidification of pksP-containing PLs (Fig. 2E). Our data suggest that the activation of calmodulin contributes to lipid-raft microdomain formation and its inhibition by the capability of DHN-melanin to sequester Ca2+ reduced microdomain formation.
Lipid-raft microdomains of RAW264.7 macrophages contain flotillin but not caveolin
The role of lipid rafts for phagosome biogenesis and their composition in the phagosomal membrane are unknown. Nevertheless, the differentiation of membrane microdomains into subclasses of lipid-raft domains and caveolae domains is widely accepted (Lingwood & Simons, 2010, Pike, 2009). In RAW264.7 macrophages, we did not detect caveolin-1 or caveolin-2 by immunofluorescence analysis or immunoblotting studies of whole cell extracts (Fig. 3A). Both caveolin-1 and caveolin-2 were detected in control HeLa cells (Fig. 3A). By contrast, flotillin-1, a marker for a different subset of lipid rafts and assumed to act as scaffolding protein that stabilizes lipid-raft microdomains (Banning, Tomasovic et al., 2011, Otto & Nichols, 2011, Stuermer, 2011), colocalized with GM1 ganglioside on phagolysosomal membranes of RAW264.7 macrophages containing pksP conidia but not melanized conidia (Fig. 3B, C). These data indicate that pksP but not wild-type conidia reside in PLs containing flotillin-dependent lipidraft microdomains in the membrane.

A) Western blot analysis for detection of caveolin in RAW264.7 and HeLa cells. αGAPDH = control. B) Detection of Flot-1 and lipid rafts in conidia-containing phagolysosomal membrane. Conidia were stained with CFW (blue), RAW264.7 macrophages for lipid rafts and Flot-1 with CTB-Alexa 647 and an αFlot-1 antibody, respectively. Scale bar = 5 μm C) Channel intensity plot (left) reflecting the CTB and αFlot-1 fluorescence signal of the phagolysosomal membrane surrounding the pksP conidium (right).
The number of pksP conidia residing in acidified PLs is drastically reduced in macrophages of Flot-1/Flot-2 double knockout mice
In mice and humans, two flotillins are known, designated flotillin-1 (Flot-1) and flotillin-2 (Flot-2) (Babuke, Ruonala et al., 2009, Frick, Bright et al., 2007). Both isoforms are integrated in lipid rafts by N-terminal hairpin structures and together with other proteins are suggested to form hetero-oligomeric complexes (Solis, Hoegg et al., 2007). Knockdown of either Flot-1 or Flot-2 in the murine macrophage-like cell line J774A.1 reduced the acidification of PLs containing pksP conidia; the effect by knockdown of Flot-1 was more pronounced (Fig. S5). Simultaneous knockdown of both Flot-1 and Flot-2 further decreased the percentage of acidified PLs containing wild-type conidia.
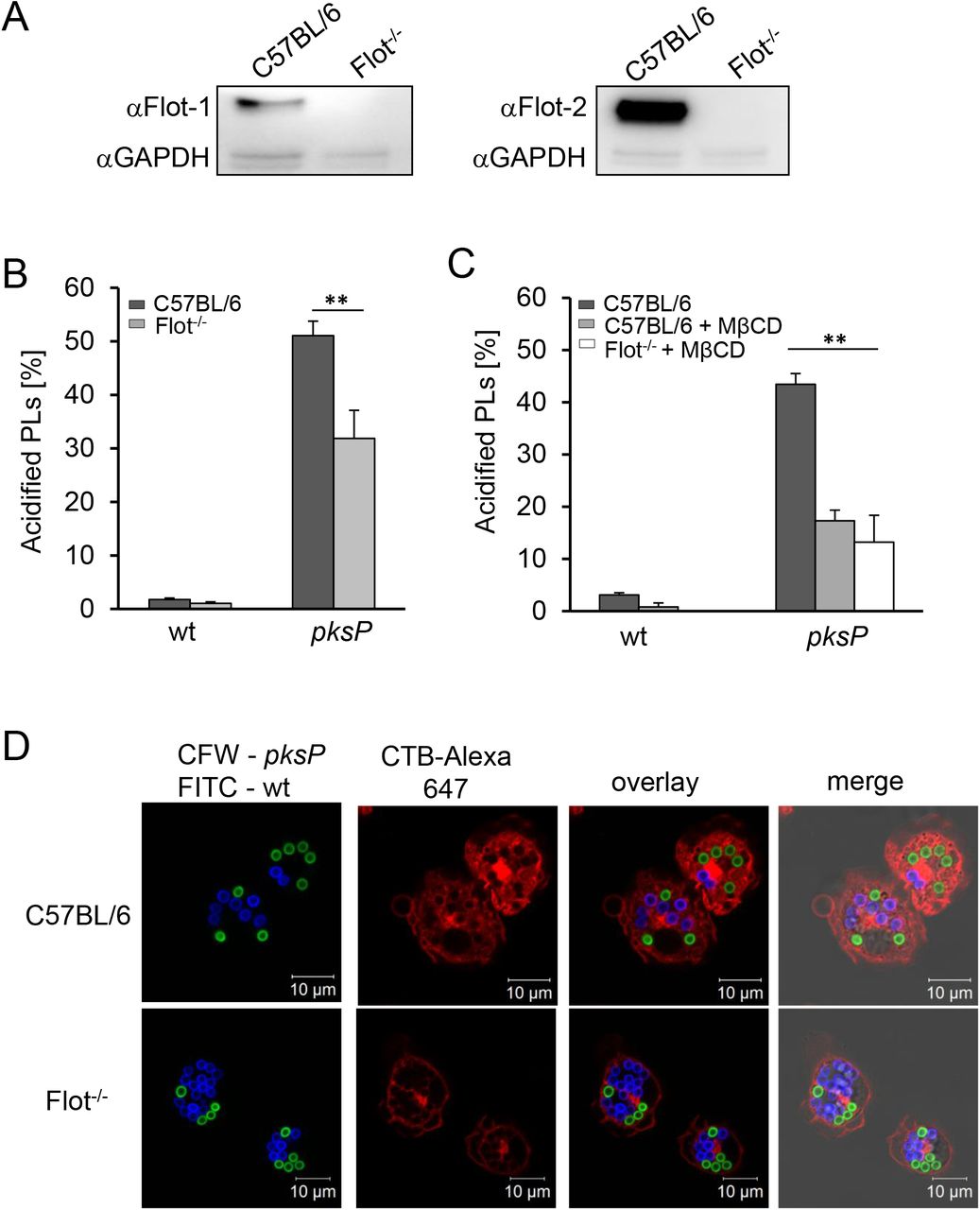
This initial finding was substantiated by the analysis of bone marrow-derived macrophages (BMDMs) from Flot-1/Flot-2 double knockout (Flot-/-) mice (Bitsikas, Riento et al., 2014) (Fig. 4A). Compared to C57BL/6 wild-type mice, BMDMs of Flot-/- mice had impaired acidification of PLs containing pksP conidia (Fig. 4B). As expected, melanized wild-type conidia containing PLs showed no significant acidification (Fig 4B).

A) Western blot analysis for detection of Flot-1 and Flot-2 in murine BMDMs of Flot-/- and wildtype mice (C57BL/6). αGAPDH = control. B) Quantification of conidia-containing acidified PLs of wild-type or Flot-/- BMDMs. Data are represented as mean ± SD; ** = p < 0.001. C) Quantification of acidified PLs in BMDMs from wild-type (C57BL/6) or Flot-/- mice. When indicated, MβCD was added. Data are represented as mean ± SD; ** = p < 0.001. D) Coincubation of BMDMs with wild-type (FITC-labeled, green) and pksP (CFW-labeled, blue) conidia. CTB-Alexa 647 stained GM1 gangliosides (red). See also Figure S5.
We also confirmed that in primary mouse BMDMs acidification of A. fumigatus conidia-containing PLs was dependent on lipid-raft microdomains. In primary macrophages, only about 5% of PLs containing wild-type conidia were acidified, and this percentage was further reduced by treatment with MβCD. Furthermore, no phagolysosomal acidification was detectable in Flot-/- macrophages treated with MβCD (Fig. 4C). The number of acidified PLs containing pksP conidia was drastically reduced by treatment with MβCD and was not further decreased in Flot-/- macrophages treated with MβCD (Fig. 4C), indicating that flotillin and cholesterol function in the same pathway to enable phagolysosomal acidification. These results were further substantiated by using Flot-/- knockout BMDMs which clearly showed reduced lipid-raft microdomains indicated by GM1 staining in the membrane of PLs containing either wild-type or pksP conidia (Fig. 4D).
Assembly of vATPase and NADPH oxidase, and phagocytosis of conidia depend on flotillin-containing lipid-raft microdomains
Previous studies reported that in maturing phagosomes, vATPase is assembled in lipid-raft microdomains (Dermine, Duclos et al., 2001, Lafourcade, Sobo et al., 2008). To proof that flotillin as a lipid-raft chaperone is in fact required for this process, the assembly of vATPase in macrophages isolated from wild-type and Flot-/- mice, was quantified (Fig. 5A). The percentage of phagolysosomal membranes showing a high degree of assembled vATPases when pksP conidia were ingested was reduced from 30 % in wild-type macrophages to about 10 % in Flot-/- macrophages, indicating the dependency of vATPase assembly on flotillins, as earlier shown for cells of the murine macrophage-like cell line J774 (Dermine et al., 2001). STED (Stimulated Emission Depletion) microscopy confirmed colocalization of Flot-1 and vATPase V1 complex (Fig. 5B) at the phagolysosomal membrane.

A) vATPase assembly in BMDMs isolated from C57BL/6 wild-type and Flot-/- mice measured by immunofluorescence. Data are represented as mean ± SD; * = p < 0.05. B) Colocalization of vATPase and Flot-1. STED microscopy of isolated PLs stained with anti-flotillin-1 and anti-vATPase V1 antibody. C) NADPH oxidase assembly in BMDMs isolated from C57BL/6 wild-type and Flot-/- mice measured by immunofluorescence. See also Figure S6. For vATPase and NADPH oxidase at least 100 intracellular conidia were evaluated for the presence of a fluorescence signal. The values represent mean ± SD of three independent experiments; D) Phagocytosis of conidia by Flot-/- BMDMs. E) Killing of conidia by Flot-/- BMDMs. The values represent mean ± SD of three independent experiments; ** = p < 0.001.
Interestingly, immunofluorescence revealed that the assembly of the NADPH oxidase complex on the phagolysosomal membrane was also affected by the absence of flotillins (Fig. S6). As shown in Fig. 5C, the percentage of PLs showing assembled NADPH oxidase complex when pksP conidia were ingested was reduced from 10 % in wild-type macrophages to 2 % in Flot-/- BMDMs, also indicating the dependency of NADPH assembly on flotillins. Taken together, deletion of flotillin genes fully phenocopied the effect of melanin on PLs. As shown in Fig. 5D, phagocytosis rates of pksP conidia were reduced in Flot-/- BMDMs. There was no effect on the phagocytosis rate of wild-type conidia, since these conidia via their DHN-melanin layer most likely already led to reduced flotillins in the membrane forming the phagocytic cup. Intracellular killing of conidia was not affected by Flot-/- BMDMs compared to wild-type BMDMs (Fig. 5E).
FLOT1 SNP results in heightened susceptibility for invasive aspergillosis in hematopoietic stem-cell transplant recipients
Our data suggested that flotillin-dependent lipid-raft microdomain formation plays a key role in immunity to infection with A. fumigatus. Therefore, we next explored the role of this pathway in humans. We screened for single nucleotide polymorphisms (SNPs) and their association with the risk of invasive aspergillosis (IA) in a cohort of hematopoietic stem-cell transplant (HSCT) recipients (Stappers, Clark et al., 2018). Whereas for the FLOT2 gene there was no SNP associated with risk of infection, remarkably, rs3094127, a SNP (T>C) in the last intron of the FLOT1 gene that is lacking in the corresponding FLOT1 gene in mice (Fig. 6A), was associated with an increased risk of IA after transplantation (Fig. 6B). The cumulative incidence of IA for donor rs309412 was 38.5% for CC (p = 0.04), 29.6% for TC (p = 0.05) and 19.6% for TT genotypes, respectively (Fig. 6B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A) Position of SNP found in human FLOT1 gene. The SNP changes T to C.
B) Cumulative incidence of IA in recipients of allogeneic hematopoietic stem-cell transplantation according to donor rs3094127 genotypes in FLOT1. Data were censored at 24 months, and relapse and death were competing events. p values are for Gray’s test.
C) Cytokines released by monocyte-derived macrophages obtained from healthy donors with different FLOT1 genotypes after stimulation with A. fumigatus conidia for 24 h.
In a multivariate model accounting for clinical variables associated with or tending towards IA in our cohort, the CC genotype at rs3094127 in FLOT1 remained an independent predictor of IA (HR 1.89; 95 % CI 1.14-5.67; p = 0.03). The FLOT1 genotypes had no impact in overall survival. To demonstrate a functional effect of the SNP in Flot-1 on myeloid cell function, we analyzed the responses of monocyte-derived macrophages isolated from healthy genotyped donors. We found that macrophages from the individuals carrying this SNP produced significantly less IL-1β and IL-6 following in vitro stimulation with A. fumigatus conidia compared to controls, whereas cytokines TNF-α and IL10 did not show significant differences (Fig. 6C).
DISCUSSION
Our study adds novel mechanistic insight on the role of lipid raft-based signaling in regulation of phagosome biogenesis and provides a new molecular virulence mechanism of melanin-induced inhibition of this process via the disruption of lipid rafts. The lipid microdomains (rafts) hypothesis was orginally proposed by Simons and Ikonen (Simons & Ikonen, 1997), imagining these lipid rafts as floating islands in the membrane (Luo, Wang et al., 2008, Triantafilou, Miyake et al., 2002). Lipid rafts are defined as small (10–200 nm) heterogeneous, highly dynamic, sterol (cholesterol), sphingolipid and protein-enriched domains that compartmentalize the cellular processes (Pike, 2006, Varshney, Yadav et al., 2016). One of the widely appreciated roles of lipid rafts is the recruitment and concentration of molecules involved in cellular signalling. The formation of a molecular cluster and their signal transduction machinery in membrane rafts leads to enhanced signalling efficiency (Triantafilou et al., 2002). It is thus not surprising that lipid rafts are required for immunity (Varshney et al., 2016). The exact role of lipid rafts and their composition are still a matter of debate. Nevertheless, the differentiation of membrane microdomains into sub-classes of lipid-raft domains and caveolae domains is widely accepted (Lingwood & Simons, 2010, Pike, 2009).
Here, we characterized lipid rafts in the phagolysosomal membrane by using a variety of marker molecules like GM1 ganglioside and cholesterol. To further specify the membrane microdomains, we assessed the localization of typical lipid raft marker proteins during PL maturation. Our findings are in accordance with studies underlining that caveolins are not coexpressed in murine macrophages (Gargalovic & Dory, 2001, Nagao, Ishii et al., 2010). Instead, our data indicate that flotillin-dependent lipid-raft membrane domains play a pivotal role in phagosome biogenesis and acidification of PLs (Lafourcade et al., 2008) and that these membrane structures are targeted by A. fumigatus conidia to interfere with the maturation of this compartment. These findings were confirmed by the observation that flotillin-1, a marker for a different subset of lipid rafts and assumed to act as scaffolding protein that stabilizes lipid rafts (Banning et al., 2011, Otto & Nichols, 2011, Stuermer, 2011), colocalized with GM1 ganglioside on phagolysosomal membranes of RAW264.7 macrophages containing pksP conidia but not melanized conidia. Further, knockdown and knockout of the flotillin genes had a major influence on the acidification of PLs and the lipid-raft assembly on the phagolysosomal membrane. Consistently, flotillins were found to be required for vATPase assembly, explaining the lack of acidification of PLs containing wild-type conidia.
Flotillins belong to a family of lipid-raft-associated integral membrane proteins. Flotillin members are ubiquitously expressed and located to non-caveolar microdomains on the cell membrane. Two flotillin members have been described, flotillin-1 and flotillin-2 (Otto & Nichols, 2011, Vieira, Correa et al., 2010). They constitutively associate with lipid rafts by acylation (a single palmitate in flotillin-1, a myristate and three palmitates in flotillin-2), oligomerization and cholesterol binding (Meister & Tikkanen, 2014). Flotillins have been long considered markers of lipid rafts because they are detergent insoluble and float in sucrose density gradients. Consequently, flotillins, either on their own or in combination with the respective other flotillin, have been implicated in numerous signalling events and pathways that are thought to be organised in lipid rafts (Babuke & Tikkanen, 2007). As shown here, by emplyoing immunostaining, BMDMs of Flot-/- mice and knockdown cell lines, the lipid rafts on the phagolysosomal membrane contain and require flotillin proteins.
An important aspect found here is that flotillin-dependent lipid-rafts form a platform for vATPase assembly and NADPH oxidase complex assembly on the phagolysosomal membrane. Further, they contribute to phagocytosis. These findings not only resulted from experiments with knockout cells but also from the colocalization of vATPase and Flot-1 on the phagolysosomal membrane. The vATPase is required for pumping protons in the lumen of the PL and thereby acidifying its content. The V1 complex of vATPase is located in the cytoplasm and contains the active site responsible for hydrolyzing ATP. The reversible binding of the V1 to the V0 complex in the phagosomal membrane regulates the activity of vATPase (Cotter et al., 2015). Little is known about the role of the vATPase and its regulation associated with the endocytic pathways of immune cells although this multiprotein complex is crucial for the antimicrobial properties of professional phagocytes, e.g., intracellular killing, digestion, and presentation of antigenic epitopes (Cotter et al., 2015, Lukacs, Rotstein et al., 1990). Genome wide knockout studies in Saccharomyces cerevisiae showed that the presence of sphingolipids strongly influences the assembly of the vATPase complex (Chung, Lester et al., 2003, Finnigan, Ryan et al., 2011). In addition to cholesterol, sphingolipids are key components of lipid rafts (Fessler & Parks, 2011, Pike, 2009, Simons & Gerl, 2010, Simons & Toomre, 2000). There is a direct link between lipidraft microdomains and regulation of the vATPase (Dhungana, Merrick et al., 2009, Foster, De Hoog et al., 2003, Lafourcade et al., 2008). Here, by using knockout BMDMs and knockdown cell lines we obtained evidence that the assembly of a functional vATPase on the phagolysosomal membrane requires flotillin-dependent lipid rafts. In line, by coinfecting macrophages with both wild-type and pksP conidia, not only different acidification patterns of PLs in the same cell occurred but also different localization patterns of the cytoplasmic V1 vATPase complex. All these effects were due to different amounts of lipid rafts in the phagolysosomal membrane. Thus, dysregulation of phagolysosomal lipid rafts by melanized conidia is not an overall mechanism of the entire cell but locally restricted to the specific phagolysosomal compartment containing a conidium. These findings correspond with previous results showing that during PL maturation the association of phagolysosomal compartments with lipid rafts governs increased acidification due to higher vATPase assembly rates (Lafourcade et al., 2008). As shown for LAP (Kyrmizi et al., 2018), the restriction of these effects to a distinct PL is most likely due to the binding of activated calmodulin to the phagolysosomal membrane. If there is free Ca2+ available for release from the phagolysosomal lumen to the cytosolic part of the phagosomal membrane, calmodulin on the cytosolic part of the membrane is activated and as a result the PL is functional, as seen here for PLs containing pksP conidia.
An interesting finding is the dependence of NADPH oxidase complex assembly on flotillin-dependent lipid rafts. It is conceivable that defects in flotillins result in immune suppression, e.g., as shown here phagocytosis was reduced in Flot-/- BMDMs or reactive oxygen species (ROS) produced by NADPH oxidase are required for induction of LAP (Martinez, Malireddi et al., 2015). This assumption is supported by the reduced phagocytosis rate of pksP conidia in Flot-/- BMDMs suggesting that lipid rafts are required as signaling platforms for recognition of conidia. By contrast, wild-type conidia did not show different phagocytosis rates because these conidia via their DHN-melanin layer most likely already disturbed flotillins in the membrane forming the phagocytic cup. In line, recently it was shown that flotillin-1 facilitates inflammatory toll-like receptor 3 signaling in human endothelial cells (Fork, Hitzel et al., 2014).
Until now, the regulation of lipid-raft formation is a matter of debate. Here, we found that inhibition of Ca2+-dependent calmodulin activity or melanin-dependent Ca2+ sequestration in the PL reduced the presence of lipid rafts on the phagolysosomal membrane. Calmodulin is a versatile Ca2+-sensor/transducer protein that modulates hundreds of enzymes, channels, transport systems, transcription factors, adaptors and other structural proteins, controlling in this manner multiple cellular functions. It can regulate target proteins in a Ca2+-dependent and Ca2+-independent manner (Villalobo, 2018). Since we found clear differences in the concentration of free Ca2+ in PLs containing pigmentless pksP conidia versus melanized conidia, this data suggests that the available free Ca2+ ions for calmodulin are reduced and thereby calmodulin activation and lipid-raft formation. This conclusion was substantiated by the finding of reduced lipid-raft formation on the phagolysosomal membrane when cells were treated with a calmodulin inhibitor. Our data well agree with our previous observation that early, transient and selective calmodulin localization was exclusively observed in PLs of melanin-deficient conidia (Kyrmizi et al., 2018).
Until now, for different pathogens such as viruses, bacteria and protozoa, it was shown that they can use host-cell lipid rafts to secure their entrance and maintenance inside target cells (Manes, del Real et al., 2003, Vieira et al., 2010). Different viruses have evolved strategies to subvert raft-associated signalling, enabling their efficient replication in immune cells, and at the same time blocking the immune response that is elicited by the target cells (Hawkes & Mak, 2006). Likewise, several bacteria interact with host lipid rafts to enter and survive inside the cell (Hawkes & Mak, 2006, Manes et al., 2003). The mechanisms that underlie this interaction are starting to be unravelled. Activation of secretion, binding, perforation of the host-cell membrane and signalling to trigger bacterial phagocytosis are involved with components of membrane microdomains (Lafont & van der Goot, 2005, Vieira et al., 2010).
Maza and colleagues (2008) investigated yeast forms of the fungal pathogen Paracoccidioides brasiliensis in the context of kinase signalling. This pathogen apparently promotes the aggregation of lipid rafts in epithelial cellls allowing fungal adhesion (Maza, Straus et al., 2008). Here, we uncovered a novel virulence mechanism. Unlike many facultative intracellular pathogens, as shown here A. fumigatus evades phagocytes by impairing lipid-raft formation in phagolysosomal membranes most likely via Ca2+ sequestration by melanized conidia. Thereby, melanized conidia prevent lipid raft-associated assembly of the signal platforms required, e.g., for assembly of vATPase and NADPH oxidase. However, alternatively it is also conceivable that incomplete or only partial fusions have occurred between endosomal and lysosomal vesicles instead of complete fusions with entire mixing of the fusion partners’ membranes (Desjardins, 1995, Haas, 2007). Since lipid rafts are also associated with functional membrane trafficking (Simons & Gerl, 2010), missing microdomains in the wild-type conidia-containing phagolysosomal membranes could be responsible for the lack of integration of lysosomal membrane constituents, which then leads to PLs devoid of lipid-raft microdomains.
Recently, we reported that DHN-melanin inhibits the activation of a noncanonical LAP autophagy pathway (Akoumianaki et al., 2016), which is regulated by NADPH oxidase and which promotes fungal killing. It was found that DHN-melanin inhibits NADPH oxidase-dependent activation of LAP by excluding the p22phox subunit from the PL. Also, NOX2-generated ROS are necessary for LC3 recruitment to phagosomes (Huang, Canadien et al., 2009). Here, we provide a model explaining that conidial melanin interferes with lipid-raft formation that is required for NADPH oxidase assembly and most likely all further processes assigned to the activity of melanin.
The importance of the lipid-raft component flotillin for pathogenicity was impressively demonstrated by the analysis of a total of 370 hematologic patients undergoing allogeneic hematopoietic stem-cell transplantation. We identified a SNP in a region of the FLOT1 gene that is not present in mice and which results in heightened susceptibility for invasive aspergillosis. Macrophages of individuals homozygous for this SNP showed reduced extracellular amounts of IL-1ß and IL-6. IL-1ß has been clearly shown to be required for defense against A. fumigatus infection (Sainz, Perez et al., 2008, Wojtowicz, Gresnigt et al., 2015) and flotillin dependent lipid rafts have been shown to affect cytokine secretion (Kay, Murray et al., 2006). These data show the functional impairment caused by the SNP and the importance of Flot-1-dependent lipid rafts for immunity against infections. In the FLOT2, gene we did not detect a SNP that could be correlated with susceptibility for invasive aspergillosis. This finding is also supported by the observation that the knockdown of either FLOT1 or FLOT2 in the murine macrophage cell line J774A.1 both reduced the acidification of PLs containing pksP conidia, but knockdown of FLOT1 had a greater effect. Until now, it is a matter of debate whether flotillins 1 and 2 are codependent in their cellular functions, or whether they can also function individually that was shown for a number of cellular effects (Bitsikas et al., 2014, Langhorst, Reuter et al., 2008, Langhorst, Solis et al., 2007, Neumann-Giesen, Fernow et al., 2007, Schneider, Rajendran et al., 2008, Stuermer, 2011, Vieira et al., 2010). Thus, it is likely that flotillins can act as heteromers but also independent of each other.
Collectively, our data provide new insight into the importance of lipid rafts for immunity against human pathogenic fungi and as a target for pathogens. Our study adds novel mechanistic insight into the regulation and formation of lipid rafts, elucidates a role for lipid raft-based signaling in regulation of phagosome biogenesis and reports a new molecular virulence mechanism via the disruption of lipid-raft microdomains.
Molecules on the fungal surface and excreted molecules belong to the first structures interacting with host cells. Although the recognition of immunological cell wall structures like β-1,3-glucans (Steele, Rapaka et al., 2005) which are more susceptible for receptors on pksP conidia (Luther et al., 2007) leads to an activation of immune cells by generation of an inflammatory response through production of chemokines and cytokines (Chai et al., 2010), it does not necessarily result in a higher activation of the endocytic pathway with accompanied acidification of phagolysosomal compartments. This was concluded here from experiments, in which the same macrophage had phagocytosed both a wild-type and a pksP conidium, which, in the same cell, ended in a neutral and acidified phagolysosome, respectively. Instead, each ingested particle determines its intracellular fate via influencing the endocytic pathway by its morphological and chemical properties, in the case of A. fumigatus conidia by the presence or absence of the DHN-melanin layer. As shown here, abolishing the phagolysosomal acidification by disturbance of vATPase assembly resulted in increased host cell damage, interestingly to the same extent for both wild-type and pksP conidia. Thus, phagolysosomal acidification strongly contributes to the effective killing of conidia (Jahn, Langfelder et al., 2002) and thereby protects to a certain extent the host cell from outgrowth of the ingested pathogen. Inhibition of phagolysosomal acidification and reduction of NADPH oxidase complex assembly by DHN-melanin allows wild-type conidia to create a favorable niche with a nearly neutral pH, which prevents activation of hydrolytic enzymes and thereby phagolysosomal digestion.
EXPERIMENTAL PROCEDURES
Cultivation of fungal strains, cell lines and infection experiments
The A. fumigatus strains used in this study, the ATCC46645 wild type and the non-pigmented pksP mutant were cultivated on Aspergillus minimal medium (AMM) agar plates as described elsewhere (Gsaller, Hortschansky et al., 2014, Jahn et al., 1997). MH-S (ATCC:CRL-2019), J774A.1 (ATCC:TIB-67), HeLa (ATCC:CCL-2), and RAW264.7 cells (ATCC:TIB-71) were cultivated in RPMI 1640 and DMEM, respectively, at 37 °C, 5 % (v/v) CO2 in a humidified chamber. A detailed protocol is provided in the Supplemental Information. The calmodulin inhibitor W7 (A3281, Sigma-Aldrich) was used in a concentration of 10 μM.
Host cell damage assay
Host cell damage caused by A. fumigatus was measured by the previously described 51Cr release assay (Filler, Swerdloff et al., 1995). A detailed protocol is provided in the Supplemental Information.
Immunofluorescence and sample preparation for CLSM and STED
See the Supplemental Information for detailed description.
Quantitation of phagolysosomal acidification and lipid raft recruitment
Prior to infection, macrophages were preloaded with 50 nM LysoTracker Red DND-99 (Life Technologies) in medium for 1 h in the absence or presence of 7.5 mM methyl-β-cyclodextrin (MβCD) (Sigma-Aldrich) or after addition of 1 μM Alexa Fluor 647-conjugated Cholera Toxin B (Life Technologies), cells were infected with A. fumigatus conidia. Then, fixed or live cells were subjected to microscopic analysis. The values represent mean ± SD of three different experiments. A detailed protocol is provided in the Supplemental Information.
Subcellular fractionation and Western blot analysis
After coincubation of conidia and immune cells, cytoplasmic and host membrane-containing cell fractions were obtained using the Qproteome Cell Compartment Kit (Qiagen) according to the manufacturer’s instructions. PLs were prepared from RAW264.7 macrophages as previously described (Akoumianaki et al., 2016). A detailed protocol is provided in the Supplemental Information.
Extraction and quantification of cholesterol
Cholesterol measurements were performed using liquid chromatography coupled to triple-quadrupole mass spectrometry (LC-MS/MS). A detailed protocol is provided in the Supplemental Information.
Calcium staining
Cells were loaded at a 1:1 ratio of Fluo-4 AM (Life Technologies) and cell medium (DMEM) 30 min before coincubation. After infection with conidia (MOI = 5) cells were coincubated for 2 h and PLs were isolated as described before.
Knockdown of Flotillin genes
J774A.1 cells were seeded with a density of 3×105 or 1×105 in 6 or 24 well plates in DMEM supplemented with 10 % (v/v) FBS and 1 % (w/v) ultraglutamine. After 24 hours the cells were transfected with 40 nM target specific siRNA (Santa Cruz Biotechnology) and 3-6 μl Lipofectamin 2000 (Life Technologies). See the Supplemental Information for detailed description.
Mouse strains and isolation of murine bone marrow-derived macrophages
Flotillin-1, flotillin-2 (Flot-1, Flot-2) double knockout (Flot-/-) mice (Bitsikas et al., 2014) were a kind gift of Ben J. Nichols (Cambridge, UK). Bone marrow cells were harvested from femurs and tibias of specific pathogen-free mice according to a procedure described elsewhere (Zhang, Goncalves et al., 2001). All animals were cared for in accordance with the European animal welfare regulation. A detailed protocol is provided in the Supplemental Information.
Human studies
A total of 370 hematologic patients undergoing allogeneic hematopoietic stem-cell transplantation at the Hospital of Santa Maria, Lisbon and Instituto Português de Oncologia (IPO), Porto, between 2009 and 2014 were enrolled in the study (Stappers et al., 2018), including 79 cases of probable/proven aspergillosis and 244 uninfected controls. The cases of invasive aspergillosis were identified and classified as ‘probable’ or ‘proven’ according to the revised standard criteria from the European Organization for Research and Treatment of Cancer/Mycology Study Group (EORTC/MSG) (De Pauw, Walsh et al., 2008). Study approval for the genetic association study was obtained from the Ethics Subcommittee for Life and Health Sciences of the University of Minho, Portugal (125/014), the Ethics Committee for Health of the Instituto Português de Oncología - Porto, Portugal (26/015), the Ethics Committee of the Lisbon Academic Medical Center, Portugal (632/014), and the National Commission for the Protection of Data, Portugal (1950/015). Approval for the functional studies on human cells was provided by Ethics Subcommittee for Life and Health Sciences of the University of Minho, Portugal (SECVS-014/2015). All individuals provided written informed consent in accordance with the Declaration of Helsinki. Genomic DNA was isolated from whole blood using the QIAcube automated system (Qiagen). Genotyping of the rs3094127 SNP in the FLOT1 gene was performed using KASPar assays (LGC Genomics) in an Applied Biosystems 7500 Fast PCR system (Thermo Fisher).
Statistical analysis
Data are expressed as mean ± SD. P values were calculated by a one-way ANOVA (Bonferroni’s post hoc test). For single comparison, p values were calculated by a two-tailed Student’s t test.
EXPANDED VIEW CONTENT
Expanded View Content includes detailed experimental procedures, six supplemental figures and two supplemental movies.
AUTHOR CONTRIBUTIONS
F.S., A.T., M.R., Z.C., H.S., S.G., A.C., M.H.G., and T.H. conducted experiments and analyzed data, A.A.B., M.T.F., G.C., C.C., A.C., J.F.L., A.C.Jr., C.E., and S.G.F. designed research and analyzed data, F.S., H.S., A.T., M.R., T.H., G.C., S.G.F. and A.A.B. wrote the paper.
ACKNOWLEDGEMENTS
We are extremely grateful to Ben J. Nichols (Cambridge, UK) for providing flotillin knockout mice. We thank Maria Straßburger for projecting mouse breeding and Anna Runtze and Muhammad Rafiq for initial experiments. We thank Matthew Blango for critically reading the manuscript. This work was supported by the excellence graduate school Jena School for Microbial Communication (JSMC) funded by the Deutsche Forschungsgemeinschaft (DFG), the International Leibniz Research School (ILRS) as part of the JSMC, the Leibniz Science Campus ‘InfectoOptics’ and the DFG-funded CRC/TR 124 ‘Pathogenic fungi and their human host - Networks of interaction - FungiNet’ (project A1 to A.A.B. and project B4 to M.T.F.) and CRC 1278 ‘Polymer-based nanoparticle libraries for targeted anti-inflammatory strategies - PolyTarget’ (project B02 to A.A.B. and Z01 to M.T.F.). C.C. and A.C were supported by the Northern Portugal Regional Operational Programme (NORTE 2020), under the Portugal 2020 Partnership Agreement, through the European Regional Development Fund (FEDER) (NORTE-01-0145-FEDER-000013), and by the Fundação para a Ciência e Tecnologia (FCT) (SFRH/BPD/96176/2013 to C.C. and IF/00735/2014 to A.C.). There is no conflict of interest for any of the authors.
REFERENCES




