

Abstract
Background Pancreatic ductal adenocarcinoma (PDAC) is extremely difficult to cure. Cancer immunotherapy has shown promising results. However, PDAC is poorly immunogenic with few effective neoantigens present, while immune suppression by myeloid-derived-suppressor-cells (MDSC) and tumor-associated-macrophages (TAM) prevents T cell activation in the tumor microenvironment (TME). To address these problems, we developed a novel concept of immunotherapy which includes the generation of memory T cells to a highly immunogenic antigen like tetanus toxoid (TT) before tumor development and reactivation of these memory T cells after tumor development in parallel with delivering TT antigens as an alternative for neoantigens into the tumor cells by an attenuated bacterium Listeria monocytogenes through in vivo infection. When combined with low doses of Gemcitabine (reduces immune suppression by MDSC and TAM) these reactivated TT-specific memory T cells are now able to destroy the Listeria-TT-infected tumor cells.
Methods Before tumor development, memory T cells were generated to a highly immunogenic recall antigen tetanus toxoid (TT) through repeated immunizations with the childhood TT vaccine (like in humans; most individuals have been exposed to during childhood vaccinations). After tumor development, the same mice (Panc-02 or KPC model) received one immunization with a high dose of Listeria-TT to deliver TT inside tumor cells, followed by multiple low doses of Listeria-TT to reactivate the memory T cells to TT. Gemcitabine was administered to reduce immune suppression. CD4, CD8 T cells, MDSC and TAM were analyzed by ELISPOT and/or flow cytometry.
Results Abundant CD4 and CD8 T cell responses producing Interferon-γ, Perforin, Granzyme B, and were observed in the spleens by ELISPOT and flow cytometry. A significant reduction was observed in tumors and metastases of the Panc-02 and KPC mice, with advanced pancreatic cancer. GEM significantly reduced the percentage of MDSC and TAM in blood and/or primary tumors, respectively.
Conclusions Our data demonstrate that delivery of recall antigen Tetanus Toxoid (TT) into tumor cells with help of attenuated Listeria through in vivo infection, in parallel with reactivating existing memory T cells to TT and low doses of GEM, could be a new type of immunotherapy to treat pancreatic cancer. This novel treatment modality of Listeria-TT and GEM warrants further investigation in the clinical setting.
Introduction
Patients with PDAC have a five-year survival of only 6%. Modern systemic therapies, such as Gemcitabine (GEM), provide some survival benefits1–4, while Gemcitabine and Abraxane or FOLFIRINOX further modestly improve survival5, 6. This underscores the need for additional and innovative approaches. Cancer immunotherapy with checkpoint inhibitors has shown promising results, however, less so for PDAC7. PDAC is poorly immunogenic because of the low mutational load and the few effective neoantigens present8, 9, and immune suppression, particularly by MDSC and TAM, prevents T cell activation in the TME10–12. Moreover, evidence suggests that naïve T cells are more prone to undergo apoptosis or are less efficiently activated than memory T cells in the TME of cancer patients and tumor-bearing mice13, 14.
To address these problems we have developed a novel treatment modality using an attenuated bacterium Listeria monocytogenes15 (Listeria) to deliver a highly immunogenic recall antigen such as tetanus toxoid (TT)(as an alternative to neoantigens) selectively into tumor cells, and to reactivate existing memory T cells against TT (generated during childhood vaccinations), which in turn destroy the now highly immunogenic infected tumor cells (SI Fig 1). These memory T cells circulate in blood for life and can be reactivated at any age. Such an approach avoids the need for naïve T cells during cancer treatment. To further improve T cell responses against TT, Listeria-TT was combined with low doses of GEM (to reduce immune suppressive MDSC and TAM populations).
We have shown that Listeria attracts and infects MDSC10 (most likely through C3bi and C1q recepors16–18), which are present in large numbers in patients and mice with cancer11, 19. However, the same MDSC are selectively attracted by the primary tumor through the production of cytokines and factors11, 20, and deliver Listeria as a Trojan horse to the TME10, 19. Once at the tumor site, Listeria spreads from MDSC into tumor cells through the cell-to-cell spread mechanism specific for Listeria21. In the TME, Listeria is protected from immune clearance through strong immune suppression, but is rapidly killed in healthy tissues that lack immune suppression. Listeria can also infect tumor cells directly22. It is through this combination of the selective attraction of MDSC by bacteria and cancer, and the strong immune suppression in the TME but not in normal tissues, why Listeria is able to selectively enter the TME and to multiply and survive, but not in normal tissues10, 19, 23, 24. Based on these results we now use Listeria as a platform for the selective delivery of anticancer agents to the TME19, 23, 25.
In the current study, we used two mouse models of pancreatic cancer, i.e. a syngeneic Panc-0226 and a transgenic KPC model27, 28. The Listeria-TT was administered through intraperitoneal (i.p.) injection, and uptake by tumor cells in vivo was verified by intravital imaging. Here, we demonstrate that the combination of Listeria-TT+GEM robustly reduced pancreatic cancer at early and advanced stages in correlation with a strong increase in CD4 and CD8 T cell responses in both Panc-02 and KPC mice. Our data provide mechanistic insight into the synergistic effects between Listeria-TT and GEM for the treatment of PDAC in mice and ultimately in patients.
Materials and Methods
Animal care
C57BL/6 mice aged 3 months were obtained from Charles River and KPC mice (LSL-p53R172/+; LSLKrasG12D; Pdx1-Cre)27 were generated in the laboratories of Dr. Steven K Libutti, Department of Surgery, and Dr. Claudia Gravekamp, Department of Microbiology and Immunology, Albert Einstein College of Medicine. C57Bl/6N-Tg(Cfms-gal4-vp16)-(UAS-eCFP)29 were maintained in the laboratory of Dr. John Condeelis. All mice were housed in the animal husbandry facility Albert Einstein College of Medicine according to the Association and Accreditation of Laboratory Animal Care (AACAC) guidelines, and kept under Bsl-2 condition as required for Listeria treatments. All mice were genotyped by Transnetyx (Cordova, TN).
Cell lines
The Panc-02 cell line (Kras-G12D mutant allele) was derived from a methylcholanthrene-induced ductal adenocarcinoma growing in a C57BL/6 female mouse30. Panc-02 cells were cultured in McCoy’s5A medium supplemented with 10% FBS, Glutamine (2 mM), non-essential amino acids, sodium pyruvate (1 mM), and Pen/Strep (100 U/ml). The Panc-02 cell line expressing Dendra-2 was developed in the laboratory of Dr. Gravekamp. Briefly, Panc-02 cells were transfected with pCDNA3.1-Dendra-2, and positive cells were selected using neomycin and FACS sorting.
Listeria and Listeria-TT
In this study, an attenuated Listeria monocytogenes (Listeria) was used as the vehicle for the delivery of TT to the TME and inside tumor cells. The Listeria plasmid pGG-34, expresses the positive regulatory factor A (prfA) and Listeriolysin O (LLO)31. prfA regulates the expression of other virulence genes, and is required for survival in vivo and in vitro. The background strain XFL-7 lacks the prFA gene, and retains the plasmid in vitro and in vivo31. The coding region for the C-terminal part of the LLO (cytolytic domain that binds cholesterol in the membranes) protein in the plasmid has been deleted, but Listeria is still able to escape the vacuole31. Mutations have been introduced into the prfA gene and the remaining LLO (expressed by the pGG34 vector), which further reduced the pathogenicity of the Listeria31. Listeria-TT was developed in our laboratory as outlined in the results section.
Infection of tumor cells in vitro
The infectivity rate of the cell lines was assessed in vitro, as described previously22. Briefly, 106 cells/ml were infected with 107 of Listeria, for 1 hr at 37°C in culture medium as described above. After the incubation with gentamicin (50 μg/ml) for 1 hr (killing all extracellular Listeria bacteria), cells were washed with PBS, lysed in water, and serial dilutions were plated onto LB agar to determine the infection rate the next day.
Evaluation of Cell Death
As described previously22, tumor cell death induced by Listeria or Listeria-TT was determined in vitro as follows. 3×103 Panc-02 cells plated in 96 well plates were infected with 107 CFU of Listeria or Listeria-TT (per well), for 3 hrs at 37°C. Gentamycin (50 μg/ml) was added until live and dead cells were counted using Trypan blue staining on the next day.
Biodistribution of Listeria-TT
C57Bl/6 mice were injected with 2×106 tumor cells as described above, and 14 days later injected once with a high dose 107 CFU of Listeria-TT. Mice were euthanized at various time points after injection of the Listeria-TT, and metastases, tumors and normal tissues were dissected, weighted, and analyzed for the number of CFU of Listeria-TT as described previously23.
Dose-limiting toxicity
C57Bl6 mice were injected i.p. with various doses of Listeria-TT, and analyzed for survival over the next 20 days as described previously23.
Tumor development
Panc-02 tumor cells were injected into the mammary fat pad (105) or orthotopically in the pancreas (106) of C57Bl/6 mice. When injected into the mammary fad pat, tumor cells metastasize to different organs (predominantly in the pancreas and liver, and less abundantly in the mesenchymal lymph nodes along the GI and in the diaphragm)19, while orthotopically injected tumor cells (only used for IVMI studies) result in a pancreatic tumor but rarely metastasize32. In KPC mice, multiple primary tumors develop spontaneously in the pancreas, which metastasizes to the liver and lungs23, 27.
Treatments
Panc-02 mice. C57Bl/6 mice were immunized i.m. twice with the “human” TT (TTvac: 0.3 μg/50 μL) vaccine (one week apart) to generate the memory T cells to TT. Subsequently, Panc-02 tumor cells (105/100 μL) were injected in mammary fat pad. When tumors were 3 mm (3-5 days after tumor cell injection) or 10 mm (10-14 days after tumor cell injection), one high dose of Listeria-TT (LM-TT) (107 CFU) was injected i.p. Three days later, the GEM treatment was started and continued for two weeks (every 3 days 1.2 mg/mouse) (6 doses in total). Concomitantly, low doses of Listeria-TT were administered daily for 2 weeks (14 doses in total). Tumor weight was determined and the number of metastases by the naked eye, as we have described previously19. KPC mice. KPC mice received the same treatment after verification of the presence of tumors and metastases by PET (3.5-8 months of age). A detailed rationale for this treatment protocol and schematic view of all treatments are shown in Fig 1.
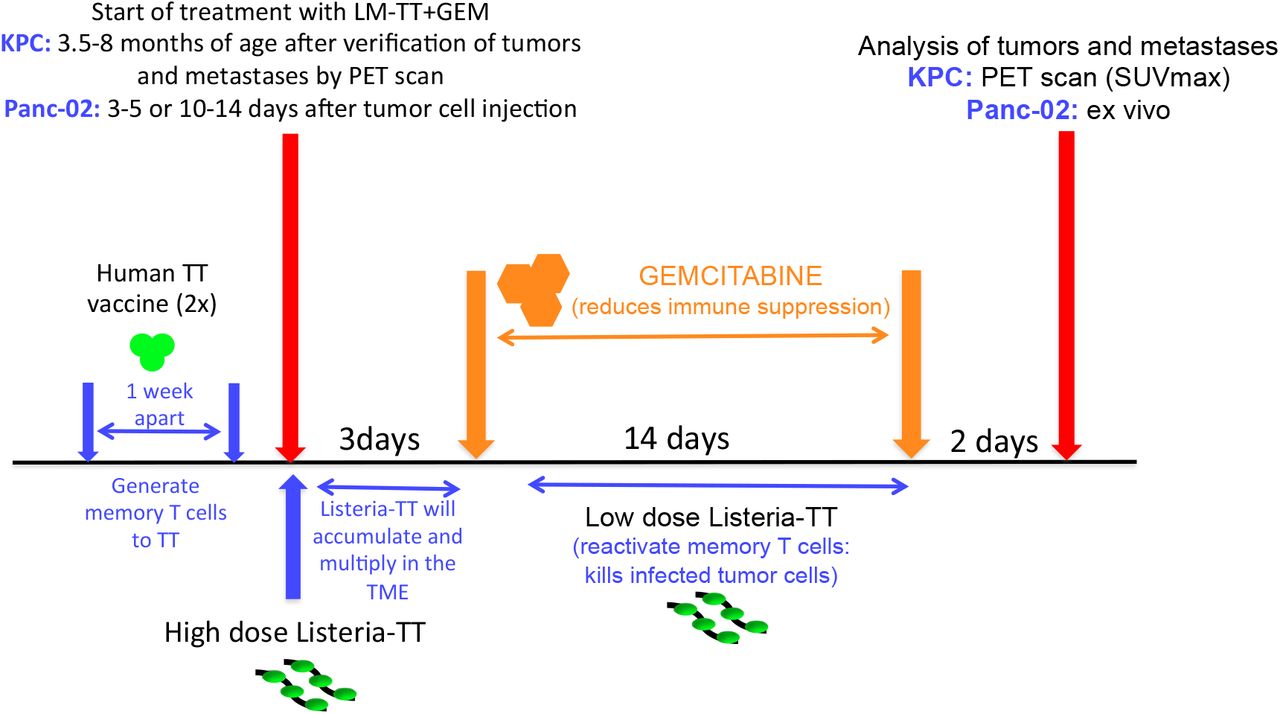
Panc-02 model: C57Bl/6 mice were immunized two times with the human TT vaccine (TTvac) one week apart to generate the memory T cells to TT similar as in humans. Subsequently, Panc-02 tumor cells (105/100 μL) were injected in the mammary fat pad. When tumors were 3 mm (3-5 days after tumor cell injectionearly pancreatic cancer) or 10 mm (day 10-14 after tumor cell injectionadvanced pancreatic cancer), one high dose of Listeria-TT (107 CFU) was injected intraperitoneally (ip). Three days later, allowing the Listeria-TT to accumulate and multiply in the TME, the treatment with GEM was started (every 3 days 1.2 mg/mouse) and continued for 14 days to eliminate MDSC and TAM (6 doses in total). At this moment MDSC are no longer required to bring the Listeria-TT to the TME. Instead, reducing the MDSC and TAM populations will improve T cell responses. Concomitantly, low doses of Listeria-TT were administered every day for 2 weeks (14 doses in total) to reactivate the T cells to TT improved by GEM. Of note is that multiple high doses of Listeria-TT induced T cell anergy but multiple low doses induced strong T cell responses to TT. All mice were euthanized 2 days after the last treatment, and analyzed for tumor weight and the number of metastases. KPC model: KPC mice of 3.5-8 months old (after having verified the presence of tumors and/or metastases by PET/CT) received the same treatments as the Panc-02 mice. The SUVmax of tumors and metastases was measured before compared to the SUVmax after treatment. LM-TT = Listeria-TT.
Intravital multiphoton imaging (IVMI)
Panc-02 tumor cells (106) expressing Dendra-2 were injected into the pancreas of transgenic mice [C57Bl/6N-Tg(Cfms-gal4-vp16)-(UAS-eCFP)]29 in which the macrophages were labeled with cyan fluorescent protein (CFP). Three weeks later, when a pancreatic tumor developed, Listeria-TT-Alexa-680 was injected i.p. Listeria-TT was labeled with an Alexa-680 dye (Listeria-TT was incubated with a rabbit anti-Listeria polyclonal antiserum (BD, Sparks, MD, cat# 223021, dilution 1:200), and anti-rabbit IgG-Alexi-680 (ThermoFisher, Rockford, IL; cat# A-21109, dilution 1:250)), on the same day as the imaging. Four hours later (when Listeria-TT had accumulated in the TME and infected tumor cells), mice were anesthetized using isofluorane (#029405, Henry Schein, Inc.). Pancreatic tumors were exposed and externalized through a small incision (~0.7 cm) through the skin and peritoneal wall and stabilized for imaging using previously published protocols33. Multiphoton intravital microscopy was performed using a custom-built two-laser multiphoton microscope34. Tumors were imaged with a femtosecond laser set to 880nm for excitation of the CFP and the green form of Dendra-2, and an optical parametric oscillator (OPO) set to 1240nm for excitation of the Alexa 680 dye. The animal was placed in a heated chamber maintained at physiological temperature during the course of imaging. Mice were monitored for heart rate, breath rate, pulse distension, breath distension and oxygen saturation using a pulse oximeter (Kent Scientific, MoseSTAT). Z-stacks of up to 45 μm of depth were acquired with a 1 μm slice interval.
PETscan
Positron Emission Tomography (PET) was performed as described previously35. Briefly, the effect of the Listeria-TT+GEM treatment on the tumor and metastases was monitored by microPET (Siemens Imaging System) before and after treatment. For this purpose, the mice were injected with positron emitter Fluorine-18 (18F)-labeled deoxyglucose (18F-FDG) and the uptake of 18F-FDG in the tumors and metastases was quantified by microPET as described previously19. Increased FDG uptake reflects increased metabolic activity of cancer cells, expressed as SUVmax.
Flow cytometry
Immune cells from spleens, blood or metastases of treated and control mice were isolated as described previously36. To identify CD8 T cells anti-CD3 and CD8 antibodies, for CD4 T cells anti-CD3 and anti-CD4 antibodies, for MDSC anti-CD11b and anti-Gr1 antibodies, and for TAM anti-CD11b and anti F4/80 antibodies were used. To detect the production of intracellular cytokines, the cells were incubated with GolgiPlug (1 ug/ml) for 6 hrs, and then treated with cytofix/cytoperm (BD Pharmingen) according to manufacturer’s instructions, for staining with antibodies to intracellular cytokines and IFNγ, Granzyme B, Perforin, IL-6, IL-10, TNFα, and other markers such as MARCO (inhibits T cell responses) CD69 (activation marker for T cells), CD80 (co-stimulatory molecule) were used. Appropriate isotype controls were used for each sample. 50,000-100,000 cells were acquired by scanning using a Fluorescence Activated Cell Sorter (Beckton and Dickinson; LSR-II, special order system), and analyzed using FlowJo 7.6 software. Cell debris and dead cells were excluded from the analysis based on scatter signals and use of Fixable Blue or Green Live/Dead Cell Stain Kit (Invitrogen). All antibodies were purchased from BD Pharmingen or eBiosciences.
ELISPOT
Spleen cells were isolated from treated Panc-02 and KPC mice and control mice for analysis by ELISPOT as described previously37. To detect T cell responses to TT, 105 spleen cells were incubated with purified TT protein (5 μg/ml). After 72 hrs, the frequency of IFNγ-producing spleen cells was measured (Pharmingen, San Diego, CA) using an ELISPOT reader (CTL Immunospot S4 analyzer, Cellular Technology, Ltd, Cleveland, OH). To determine the frequency of IFNγ-producing CD4 and CD8 T cells, spleen cells were depleted for CD4 and CD8 T cells using magnetic bead depletion techniques according to the manufacturer’s instructions (Miltenyi Biotech, Auburn, CA). All antibodies were purchased from BD Pharmingen.
Western Blotting
Expression of TT protein in tumor cells was analyzed by western blotting. Briefly, cells were lysed in RIPA buffer containing protease inhibitors, and proteins were separated on 4-12% gradient SDS-polyacrylamide gel, and then electro-transferred to PVDF filters membrane. Membranes were incubated with rabbit anti-Myc IgG antibody (Cell Signaling Technology, Danvers, MA), followed by horse-radish peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary antibodies (Santa Cruz, Dallas, TX) and detection was obtained using a chemiluminescence detection kit (Thermo scientific, Rockford, IL). Antibody to beta-actin was used to ensure equal loading.
Statistical analysis
To statistically analyze the effects of Listeria-TT+GEM on the growth of metastases and tumors or on immune responses in both mouse models Panc-02 and KPC, the Mann-Whitney was used. *p<0.05, **<0.01, ***<0.001, ****<0.0001. Values p<0.05 were considered statistically significant.
Results
Generation and characterization of Listeria-TT
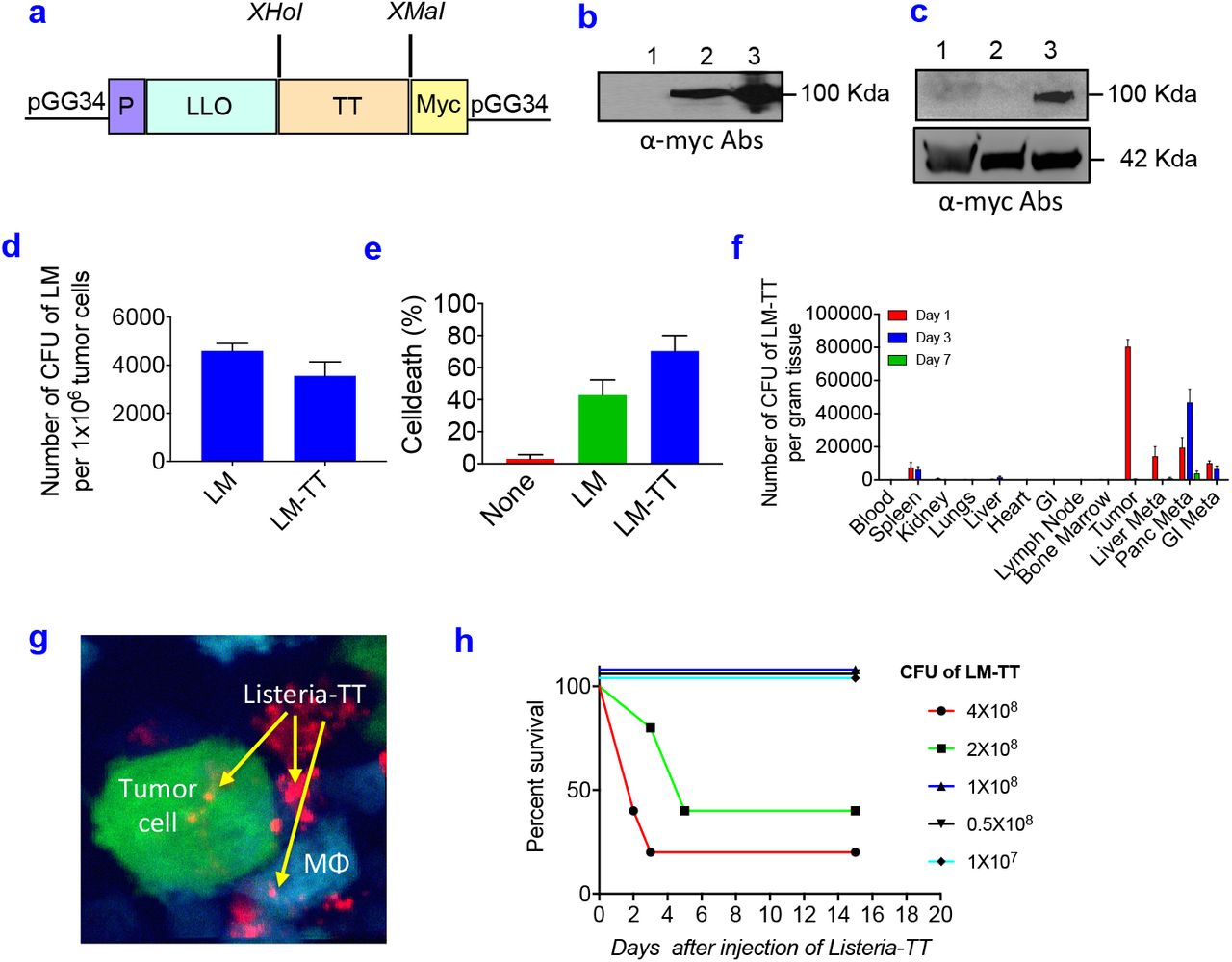
The Listeria-TT vaccine was developed as follows. The TT856-1313 (52 kDa) was cloned as a fusion-protein with a truncated Listeriolysin O (LLO)(48 kDa) in the Listeria vector (pGG34) under the control of the LLO promoter (P), and a myc sequence for detection of the TT protein was included (Fig 2a). Secretion of LLO-TT856-1313 protein by the Listeria-based vaccine was detected in the culture medium by western blotting using anti-myc antibodies (Fig 2b). Infection of Panc-02 tumor cells with Listeria-TT856-1313 resulted in the expression of TT protein in the tumor cells (Fig 2c).
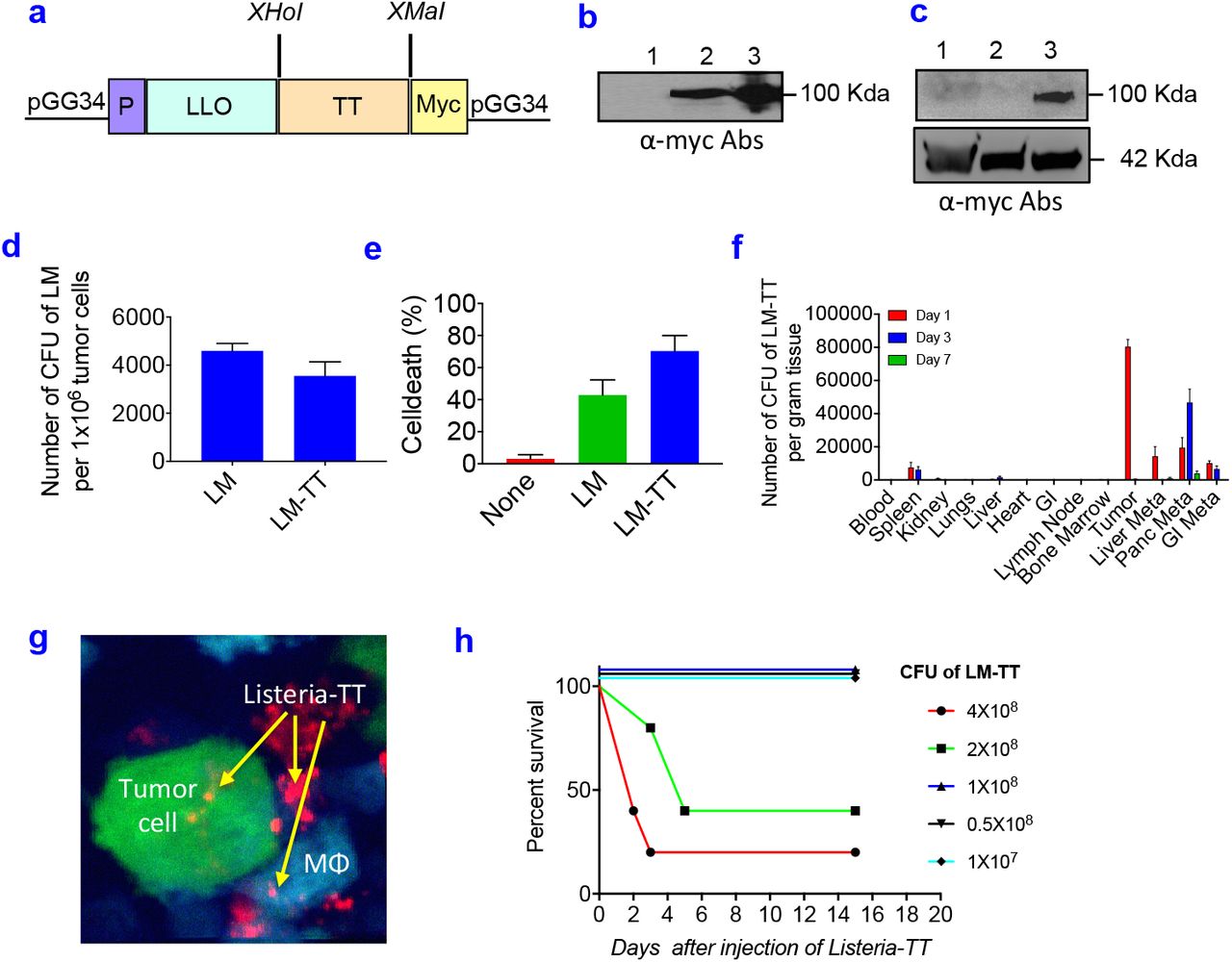
(a) Listeria-TT construct. A non-toxic fragment of the C-terminus of TT cDNA (aa position 856-1313) was cloned as a fusion protein with a truncated non-cytolytic ListeriolysinO (LLO) in the Listeria plasmid pGG34, under the control of the LLO promoter (P). A myc tag has been included for the detection of TT protein. (b) Detection of TT protein in culture supernatant. Anti-myc antibodies were used. Lane 1: neg control (medium); Lane 2: supernatant of Listeria-TT culture; Lane 3: pellet of Listeria-TT culture. (c) Detection of TT protein in tumor cells. Lane 1: Panc-02 tumor cells; Lane 2: Panc-02 tumor cells infected with Listeria alone; Lane 3: Panc-02 tumor cells infected with Listeria-TT. (d) Panc-02 tumor cells infected with Listeria-TT. (e) Panc-02 tumor cells killed by Listeria-TT. (f) Biodistribution of Listeria-TT in Panc-02 model. n=3 mice per time point. (g) Listeria-TT infecting tumor cells and macrophages (Mϕ) in Panc-02-dendra-2 tumors of live mice, by intravital imaging. Tumor cells are green, macrophages blue, and Listeria-TT red. (h) Dose-limiting toxicity study of Listeria-TT in tumor-naive C57Bl6 mice. Survival was analyzed over the next 20 days. n=5 mice per group. In figure d, e and f, the error bars represent SEM. Mann-Whitney Mann-Whitney *p <0.05, **p<0.01, ***p<0.001, ****p<0.0001. LM-TT = Listeria-TT.
To test whether Listeria-TT remained functional as a delivery platform for anticancer agents, we determined the infection rate of pancreatic tumor cells with Listeria-TT in comparison to Listeria alone. For this purpose, Panc-02 tumor cells were cultured with Listeria or Listeria-TT, lysed in water and plated on agar, as described previously19. Both abundantly infected the Panc-02 cells (Fig 2d). We also determined whether Listeria-TT was able to kill Panc-02 cells, as described previously22. As shown in Fig 2e, Listeria-TT abundantly killed the Panc-02 cells.
TT is highly immunogenic and may lead to faster elimination of Listeria-TT than Listeria alone and consequently may prevent accumulation in the TME. One high dose of Listeria-TT was injected i.p. into Panc-02 mice and the number of CFU of Listeria-TT was measured in all tissues (tumors, metastases, and normal tissues) at different time points. As shown in Fig 2f, Listeria-TT robustly accumulated in tumors and metastases, but hardly in normal tissues. Interestingly, when injected intravenously Listeria hardly reached the tumors and metastases (SI Fig 2). This was also found by others in a murine colon carcinoma model CT2638. It is most likely that the effect of immune suppression in the peritoneal cavity generated through tumors and metastases is responsible for the preferential colonization of Listeria in the TME, and that immune suppression in blood is less potent.
To confirm that Listeria was taken up by tumor cells within the TME of live mice, we injected Listeria, labeled ex vivo with Alexa-680 dye, into the peritoneal cavity of transgenic mice whose macrophages express cyan fluorescent protein [C57Bl/6N-Tg(Cfms-gal4-vp16)-(UAS-eCFP)]29 and which were bearing orthotopic Dendra-2 labeled pancreatic tumors (Panc-02-Dendra). We then imaged these tumors using high-resolution intravital multiphoton microscopy (IVMI), and showed individual Listeria inside tumor cells and macrophages (Fig 2g).
Generation of memory CD4 and CD8 T cells to TT before tumor development and reactivating these memory T cells after tumor development
First, we generated TT-specific memory CD4 and CD8 T cell responses through two immunizations with the human TT vaccine (TTvac) before tumor development (Fig 3a). Subsequently, the Panc-02 tumor cells were injected in the mammary fat pad of mice with the existing memory T cells to TT. Once the Panc-02 tumors were 10 mm in diameter, the treatment was started with one high dose of Listeria-TT injected followed by multiple low doses of Gemcitabine, concomitantly with low doses of Listeria-TT over a period of two weeks as outlined in Fig 1.

(a) Generation of CD4 and CD8 memory T cells to TT mice before tumor development through immunizations with the human TT vaccine (TTvac). (b) Reactivation of CD4 and CD8 T cell responses to TT after tumor development through a complete treatment cycle with Listeria-TT+GEM. T cells producing IFNγ before and after tumor development were analyzed in a restimulation assay with TT protein by ELISPOT. Representative of 2 experiments. n=3 mice per group (pooled). (c) GEM reduces MDSC populations in blood and primary tumors, and TAM in primary tumors. MDSC (CD11b+Gr1+) and TAM (CD11b+F4/80+) were analyzed after a full treatment cycle with TTvac, and Listeria-TT plus GEM as described in Fig 1. Representative of 2 experiments. n=3 mice per group (pooled). Mann-Whitney *p<0.05, ***p<0.001 and *****p<0.0001.
To evaluate recall of the memory T cell responses to TT after tumor development, spleen cells were isolated from the tumor-bearing mice (after the complete treatment cycle), restimulated with TT protein in vitro, and then analyzed for memory T cell responses producing IFNγ by ELISPOT. Depletion of CD4 and CD8 T cells from the total spleen population showed significant reduction in the production of IFNγ of mice that received Listeria-TT+GEM or Listeria-TT alone (Mann-Whitney p<0.05 and p<0.001), indicating CD4 and CD8 memory T cell responses to the TT protein, while no T cell responses were observed in the treatment groups that received GEM or saline only (Fig 3b).
We also analyzed T cell responses directly in vivo in mice with the Panc-02 tumor-bearing mice after a full treatment cycle with TTvac, Listeria-TT+GEM, by flow cytometry, without restimulation in vitro. Our goal was to analyze if a complete treatment cycle with Listeria-TT+GEM could reduce the immune suppression in the TME and activate T cells in vivo. T cell responses (CD69, Granzyme B, Perforin, and IFNγ) were strongly improved in the spleen (pooled) of mice treated with Listeria-TT+GEM compared to all control groups (Table 1a and SI Fig 3ab). Not only did both CD8 and CD4 T cells produce high levels of Granzyme B, Perforin, and/or IFNγ, but CD4 T cells also demonstrated an even stronger response than the CD8 T cells. Moreover, Listeria-TT or GEM alone also increased the production of Perforin in CD4 and CD8 T cells compared to the saline treatments.
Listeria-TT and GEM increases CD4 and CD8 T cell responses in vivo in Panc-02 mice
Similar results were found in the spleens of KPC mice by flow cytometry (which received the same treatment as the Panc-02 mice). CD4 and CD8 T cell responses were robustly improved in mice that received Listeria-TT+GEM compared to all control groups, i.e. the percentage of CD4 and CD8 T cells producing Granzyme B, Perforin, and/or IFNγ, or expressing CD69, was strongly increased in mice that received Listeria-TT+GEM compared to all control groups (Table 1b).
Listeria-TT and GEM increase CD4 and CD8 T cell responses in vivo in KPC mice
Finally, in a pilot study using RNAseq we found that CD3/4/8/28, Granzymes and Perforin were abundantly upregulated in pancreatic tumors of KPC mice that received TTvac, Listeria-TT+GEM compared to untreated KPC mice (SI Fig 4a). These results support the in vivo and in vitro analysis of the T cell responses to Listeria-TT+GEM (Fig 3b, Table 1ab).
Listeria-TT and GEM alter the TME
Panc-02 tumor bearing mice received a full treatment cycle with Listeria-TT+GEM as described in Fig 1. Here, we show that GEM reduced the MDSC population in blood by 80-90% and in primary tumors by 50% compared to the saline group (Fig 3c), and the TAM population by 67% in primary tumors compared to the saline group (Fig 3c).
Moreover, mice treated with Listeria-TT+GEM showed a decrease in the percentage of MDSC (Table 2a and SI Fig 5a) and TAM (Table 2b and SI Fig 5b) secreting IL-10, IL-6 or expressing MARCO (all involved in T cell inhibition) in the pancreatic metastases, but increased percentages of MDSC and TAM producing TNFα (involved in tumor cell killing) and/or CD80 (involved in T cell activation), compared to the controls. A similar effect on cytokines was shown by Listeria-TT alone but less abundantly.
Listeria-TT and GEM reduce immune suppressive function of MDSC (CD11b+Gr1+) in metastases of Panc-02 mice
Listeria-TT and GEM reduce immune suppressive function of TAM (CD11b+F4/80+) in metastases of Panc-02 mice
Reducing early and advanced pancreatic cancer in Panc-02 and KPC mice by Listeria-TT and GEM
Here we tested Listeria-TT+GEM therapeutically in Panc-02 mice with early and advanced pancreatic cancer as outlined in Fig 1. We found an almost complete elimination of early stage pancreatic cancer (Fig 4a), and a significant reduction in primary (85%; p=0.0001) and metastatic tumor burden (89%; p=0.0001) of advanced pancreatic cancer compared to the saline group in the Panc-02 model (Fig 4b and d).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
(a) Early pancreatic cancer (Panc-02). C57Bl/6 mice received a full treatment cycle with TTvac, Listeria-TT+GEM as described in Fig 1; the combination therapy was started 3 days after tumor cell injection. (b) Advanced pancreatic cancer (Panc-02). C57Bl/6 mice received a full treatment cycle with TTvac, Listeria-TT+GEM as described in Fig 1; the combination therapy was started 10-14 days after tumor cell injection. (c) Advanced pancreatic cancer (KPC). KPC mice received a full treatment cycle with TTvac, Listeria-TT+GEM as described in Fig 1; the combination therapy was started at the age of 3-5.5 months, after tumors and metastases were verified through PETscan. Tumors and metastases were measured (SUVmax) before and after the treatment period. (d) Example of pancreatic tumors (yellow circles) in the different treatment groups of the Panc-02 mice. (e) Example of the pancreatic tumors (yellow circles) in the different treatment groups of the KPC mice. In abc, n=5 mice per group (average of two experiments)(controls Saline, GEM, and LM-TT in c, n=2-3 mice). Mann-Whitney Mann-Whitney *p <0.05, **p<0.01, ***p<0.001, ****p<0.0001.
We also tested Listeria-TT+GEM therapeutically in KPC mice with advanced pancreatic cancer (3.5-8 months old). A significant reduction in the SUVmax of PDAC tumors (SUVmax 1.9 to 0.4) and liver metastases (SUVmax 2.1 to 0.3) was observed in 5 out of the 9 mice (these 5 mice were 3-5.5 months of age at the start of treatment) (Fig 4c and e). The PDAC tumors reduced by 80% (p=0.0020) and the liver metastases by 87% (p=0.03). Finally, 4 out of the 9 mice died during or shortly after treatment and could not be rescued because their extensive tumor burden prior treatment initiation (KPC mice were 8 months at the start of treatment).
The SUVmax of tumors and metastases slightly increased or decreased in the Saline and GEM control groups, and considerably decreased (50%) in the LM-TT control group but this was statistically not significant (Fig 4c).
Safety of Listeria-TT and GEM
To determine the safety of Listeria-TT+GEM, a dose-limiting toxicity study was performed as described previously23. Briefly, increasing doses of Listeria-TT were injected in C57Bl/6 mice without cancer and the survival time was determined over the course of the next 20 days. As shown in Fig 2h, the LD50 of Listeria-TT was 2×108 CFU. In our treatment studies we used 107 Listeria-TT, which is far below the LD50.
We also analyzed potential toxicity by pathological examination, 2 days after a complete treatment cycle with Listeria-TT+GEM in the Panc-02 mice as outlined in Fig 1. Again, no significant pathological effects of Listeria-TT+GEM were observed (a mild increase in leucocytes was observed in the spleen, liver and lungs) (SI Table 1a).
Finally, we analyzed liver functions of C57Bl/6 mice that received the full treatment cycle compared to the saline group (untreated). A small increase in the aminotransferase (AST) level in plasma from Listeria-TT+GEM-treated mice compared to untreated mice (107 to 137 U/L) was observed, while alanine aminotransferase (ALT) levels in Listeria-TT+GEM-treated mice were similar compared to untreated mice (15 and 16 U/L) (SI Table 1b).
Discussion
Although success has been reported with cancer immunotherapy, its utility in PDAC has been underwhelming, due to the low immunogenicity of the tumor, strong immune suppression in the TME, and less efficient activation of naïve T cells in the TME8, 9, 11–14. We addressed these problems by using a new concept of a Listeria-based immunotherapy, which includes delivery of a highly immunogenic foreign TT antigen selectively to the TME and into tumor cells through in vivo infection, by reducing immune suppression with GEM, and by avoiding the need of naïve T cells during treatment through reactivation of existing memory T cells generated during childhood.
Using the childhood TT vaccine (TTvac), memory CD4 and CD8 T cells were generated to TT before tumor development and reactivated with Listeria-TT+GEM after tumor development. This resulted in strong CD4 and CD8 T cell responses (producing Perforin, Granzyme B, and IFNγ) in the spleens of tumor-bearing Panc-02 and KPC mice. Also, RNAseq analysis showed that CD3/4/8/28, Granzymes and Perforin were abundantly upregulated in pancreatic tumors of KPC mice by TTvac, Listeria-TT+GEM compared to the untreated KPC mice (SI Fig 4a), in correlation with increased expression of cell death pathways such as Casp 1,3,4,6,8,12, Fas, and tumor necrosis factor (TNF) (SI Fig 4a). These cell death pathways are potentially activated by Perforin and Granzyme B. Moreover, RNAseq analyses demonstrated that Listeria-TT+GEM also up regulated gene pathways involved in metastatic behavior, such as Cadherins (3, 5, 6, 10, 11, 13, 15, 17), and Claudins (2, 4, 5, 6, 7, 8, 10, 20) (SI Fig 4b). Both genes are involved in adherence of cells and tight junctions. Low expression of these genes is associated with strong metastatic behavior of tumors, and high expression with poor metastatic behavior39, 40.
We demonstrated that early treatment of Panc-02 mice with Listeria-TT+GEM reduced the pancreatic tumors and metastases almost completely, i.e. 9 out of 10 mice were free of cancer, which was not possible with Listeria-TT or GEM alone. Even when the treatment was started in Panc-02 and KPC mice (12-20 weeks of age) with advanced pancreatic cancer, a strong reduction in their tumors and metastases was observed.
Listeria-based immunotherapies has also been reported by others. Keenan et al. demonstrated improved significant survival and delayed the PanIN progression in KPC mice with early PDAC (4-6 weeks of age), when immunized with Listeria (Listeria double deleted ActA and InlB, both involved in spread of Listeria)(LADD) engineered to express the Kras12GD mutation, and combined with anti-CD25 Abs and cyclophosphamide41. Another interesting study recently published includes 4T1 and CT26 mice (using the flank model), immunized with LADD expressing a viral antigen AH1, which was also expressed by their tumors. This stimulated CX3CR1+CD62L-KLRG1+PD-1lo tumor-specific CD8 T cells migrating into the AH1-expressing tumors and changed the TME by reducing Tregs and changing M2 macrophages into M1 macrophages38. In addition, mouse models of cervical, breast, and skin cancer has been successfully treated with Listeria expressing E7, Her2/neu, Flk-1, CD105, Mage-b, ISG15, and HMW-MAA42.
Of note is that the concept of Listeria used by others is different from our concept. Listeria used by others is based on the delivery of tumor-specific antigens into dendritic cells to stimulate naïve and memory T cells which are naturally expressed by the tumors, while our concept is based on colonization and changing the TME by Listeria, and using Listeria to deliver highly immunogenic recall antigens inside tumor cells through in vivo infection, and to reactivate existing memory T cells to the recall antigens now also expressed by the tumors.
Listeria has also been tested in clinical trials in patients with various cancers, including pancreatic cancer, showing safety and tumor-specific T cell responses and improved survival (for a review see Forbes et al)43.
Of note is that Listeria does not infect all tumor cells. However, we have shown in a previous study that Listeria induces immunogenic tumor cell death through ROS and that T cells were activated to multiple (if not all) TAA expressed by tumors and metastases in a metastatic breast tumor model14. This resulted in a significant reduction in the number of metastases and tumors. Based on this earlier study and results from this study (Listeria-TT abundantly killing Panc-02 tumor cells Fig 2e), it is likely that the TAA-specific T cells will destroy the non-infected tumor cells. However, this needs to be further analyzed.
Elderly cancer patients react less efficiently to vaccines than young adults44, 45, which is also true for cancer immunotherapy36, 46. The cause for this differential response is mainly due to lack of naïve T cells at older age47. We believe that our novel approach of reactivating memory T cells generated during childhood (when plenty naïve T cells are available), avoids the need for naïve T cells at older age, and thus potentially making this therapeutic strategy more effective for patients of an older age. However, this belief still needs to be validated.
Conception and Design
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, Z. Yuan, S.K. Libutti, C. Gravekamp
Development of Methodology
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, Z. Yuan, S.K. Libutti, C. Gravekamp
Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc)
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, A. Jahangir, A. Patel, K. Meena, R. Alves Da Silva, A. Beck, L. Tesfa, W. Koba, Z. Yuan, S.K. Libutti, C. Gravekamp
Analysis and interpretation of data (e.g statistical analysis, biostatistics, computational analysis)
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, A. Jahangir, A. Patel, K. Meena, R. Alves Da Silva, A. Beck, L. Tesfa, W. Koba, J. Chuy, Z. Yuan, S.K. Libutti, C. Gravekamp
Writing, Review and/or revision of the manuscript
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, Z. Yuan, S.K. Libutti, C. Gravekamp
Administrative, technical or material support (i.e., reporting or organizing data, constructing data bases)
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, Z. Yuan, S.K. Libutti, C. Gravekamp
Study Supervision
B.C. Selvanesan, D. Chandra, W. Quispe-Tintaya, Z. Yuan, S.K. Libutti, C. Gravekamp
Ethics approval and consent to participate
All in vivo experiments were approved by the NCI/NIH Animal Care and Use Committee (ACUC).
Competing interests
Dr. Gravekamp has ownership interest in a patent application filed about the Listeria-recall antigen concept (96700/2230). No competing interests were disclosed by the other authors.
Acknowledgements
This work was supported by the Pancreatic Cancer Action Network (PCAN) 422247, a private donation of Janet and Marty Spatz, NCI Administrative Supplement 3P30CA013330-44S3, and NCI cancer center support P30CA013330 (Flow Cytometry Core, Pathology Core, MicroPET Core, Genetics Core).
Abbreviations
- ALT
- : alanine aminotransferase
- AST
- : aminotransferase
- CFP
- : cyan fluorescent protein
- CFU
- : colony-forming unit
- CTL
- : cytotoxic T lymphocyte
- CD
- : cluster differentiation
- DLT
- : dose-limiting toxicity
- 18F-FDG
- : Fluorine-18 (18F)-labeled deoxyglucose
- FBS
- : fetal bovine serum
- Flk
- : Fms-like tyrosine kinase
- GEM
- : gemcitabine
- HMW-MAA
- : high molecular weight melanoma-associated antigen
- HRP
- : horse-radish peroxidase
- IFN
- : interferon
- IL
- : interleukin
- ISG
- : interferon-stimulated gene
- IVMI
- : intravital multiphoton imaging
- KLRG
- : killer cell lectin-like receptor subfamily G
- KPC
- : Kras, p53, Cre
- LADD
- : Listeria double deleted
- LLO
- : Listeriolysin O
- LM
- : Listeria monocytogenes
- Mage
- : melanoma-associated antigen
- MARCO
- : macrophage receptor with collagenous structure
- MDSC
- : myeloid-derived suppressor cells
- OPO
- : optical parametric oscillator
- PD
- : programmed cell death
- PET
- : positron emission tomography
- PDAC
- : pancreatic ductal adenocarcinoma
- prfA
- : positive regulatory factor
- SDS
- : sodium dodecyl sulfate
- SUV
- : standard uptake value
- TAM
- : tumor-associated macrophages
- TME
- : tumor microenvironment
- TT
- : tetanus toxoid
- TNF
- : tumor necrosis factor.
References




