

Abstract
The receptor for hyaluronic acid-mediated motility (RHAMM) is upregulated in various cancers. We previously screened genes upregulated in human hepatocellular carcinomas for their metastatic function in a mouse model of pancreatic neuroendocrine tumor (PNET) and identified that human RHAMMB promoted liver metastasis. It was unknown whether RHAMMB is upregulated in pancreatic cancer or contributes to its progression. In this study, we found that RHAMM protein was frequently upregulated in human PNETs. We investigated alternative splicing isoforms, RHAMMA and RHAMMB, by RNA-Seq analysis of primary PNETs and liver metastases. RHAMMB, but not RHAMMA, was significantly upregulated in liver metastases. RHAMMB was crucial for in vivo metastatic capacity of mouse and human PNETs. RHAMMA, carrying an extra 15-amino acid-stretch, did not promote metastasis in spontaneous and experimental metastasis mouse models. Moreover, RHAMMB was substantially higher than RHAMMA in pancreatic ductal adenocarcinoma (PDAC). RHAMMB, but not RHAMMA, correlated with both higher EGFR expression and poorer survival of PDAC patients. Knockdown of EGFR abolished RHAMMB-driven PNET metastasis. Altogether, our findings suggest a clinically relevant function of RHAMMB, but not RHAMMA, in promoting PNET metastasis in part through EGFR signaling. RHAMMB can thus serve as a prognostic factor for pancreatic cancer.
Main text
Metastasis accounts for 90 percent of cancer deaths. We developed a mouse model of well-defined multistage tumorigenesis: RIP-Tag; RIP-tva to identify metastatic factors [1]. We identified that the receptor for hyaluronic acid (HA)-mediated motility, isoform B (RHAMMB), significantly promotes liver metastasis of pancreatic neuroendocrine tumors (PNET) in RIP-Tag; RIP-tva mouse models [2]. Expression of RHAMM is restricted in normal adult tissues, but is upregulated in cancers [3, 4]. Increased production of glycosaminoglycan, HA, is correlated with increased migration and invasion in aggressive cancers [5]. CD44 and RHAMM are two major HA receptors. The roles of CD44 isoforms in cancer have been studied extensively, but the functions of RHAMM isoforms in tumorigenesis are less clear. RHAMM encodes 18 exons and alternative splicing generates different isoforms. RHAMMA includes all exons and RHAMMB lacks exon 4 (Fig. 1A). Here we aimed to determine the clinical relevance of RHAMMA and RHAMMB isoforms and their functions in pancreatic cancer.

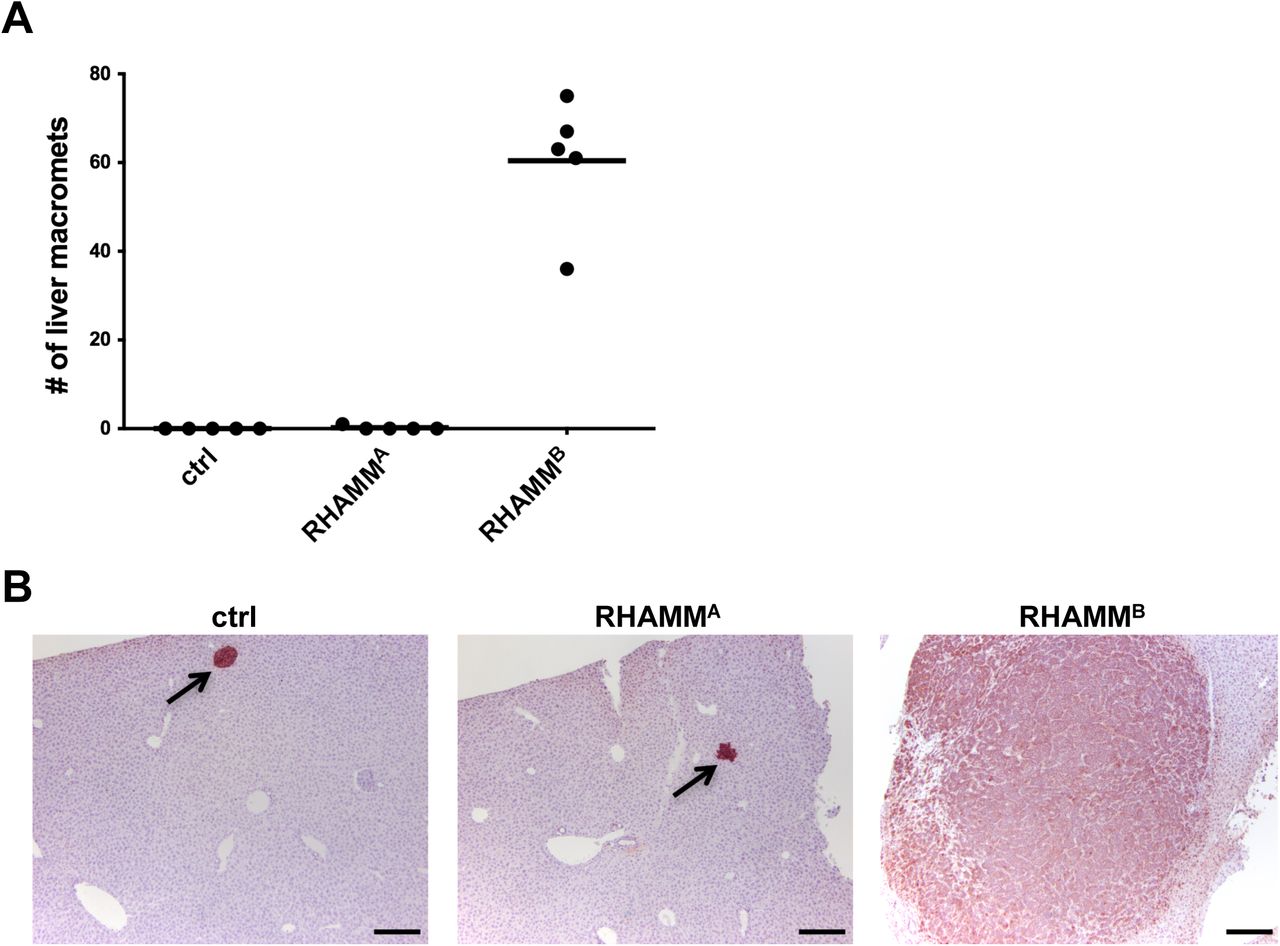
RHAMMB, but not RHAMMA, is upregulated in human PNETs and promotes liver metastasis of mouse PNET cells. (A) Diagram of RHAMMA and RHAMMB proteins. (B) RHAMM is upregulated in 54 of 83 cases (65%) of human PNETs in immunohistochemical staining. Left: Normal pancreas with islets in dashed circle. Middle: RHAMM negative PNET. Right: RHAMM positive PNET. Original magnification: 20X. Scale bar, 50 µm. (C)RNA-seq analysis showed that RHAMMB is significantly upregulated compared to RHAMMA in primary human PNETs and liver metastases. The p value was calculated using two-way ANOVA followed by Tukey’s test. **: p < 0.0001, *: p < 0.05. Error bars represent standard error of mean. (D) Western blot analysis of human RHAMM in mouse N134 cell line (control), N134-RHAMMA cells, and N134-RHAMMB cells. (E) A total of 1 million N134 cells, N134-RHAMMA cells, or N134-RHAMMB cells were injected into the tail vein of NSG mice (n = 5 for each group). Five weeks later, the recipient mice were euthanized to survey for metastatic sites and incidence. Representative liver photos were shown.
RHAMMB, but not RHAMMA, is upregulated in human PNET liver metastases
To investigate RHAMM expression in human PNETs, a tissue microarray consisting of 83 PNETs was immunostained for RHAMM. RHAMM was not detectable in the normal pancreas, while 54 of 83 (65%) PNETs exhibited cytoplasmic staining using an antibody that recognizes common region in RHAMM isoforms (Fig. 1B). Because isoform-specific RHAMM antibodies were not available, we investigated the mRNA levels of RHAMMA and RHAMMB by performing RNA-Seq analysis on 27 primary PNETs and 12 liver metastases, using 89 human islets from Gene Expression Omnibus database for comparison. Consistent with our immunohistochemical data, normal islets had very low RHAMMA and RHAMMB mRNA (Fig. 1C). RHAMMB was significantly higher than RHAMMA in both primary and metastatic PNETs, suggesting that RHAMMB was the predominant isoform naturally expressed in PNETs. Although RHAMMA levels in primary tumors were significantly higher than those in normal islets (p = 0.0002), RHAMMA levels in metastatic tumors were not significantly higher than those in normal islets (p = 0.8928). The mRNAs of RHAMMA and RHAMMB were readily detectable in additional primary PNETs and metastases by RT-qPCR using isoform-specific primers (Fig. S1A-B).

RT-qPCR analysis of RHAMMA (A) and RHAMMB (B) in 9 primary human PNETs and 3 metastatic PNETs from the livers.
We compared metastatic potential of RHAMMA to RHAMMB in RIP-Tag; RIP-tva models of spontaneous metastasis and tail vein assays [1, 2]. In contrast to RHAMMB, RHAMMA did not promote spontaneous metastasis (Table S1). Then, we generated N134 cells overexpressing RHAMMA (N134-RHAMMA). N134 is a cell line derived from a PNET of RIP-Tag; RIP-tva mouse [1]. Although there were more RHAMMA than RHAMMB for unknown reasons (Fig. 1D), only one visible tumor was found in 5 immunodeficient NOD/scid-IL2Rgc knockout (NSG) mice receiving N134-RHAMMA cells after 5 weeks, while all 5 mice receiving N134-RHAMMB cells developed large liver metastases within 5 weeks (Fig. 1E and Fig. S2A). To detect micrometastases, we performed immunostaining for synaptophysin, a neuroendocrine marker. Mice receiving N134 cells and N134-RHAMMA cells had an average of 1.8 and 0.6 liver micrometastases, respectively (Fig. S2B). These data suggest that the unique 15-amino acid-stretch, ESQKNDKDLKILEKE, which is present in RHAMMA but not in RHAMMB, inhibited the metastatic function of RHAMM.
Impact of genes on lymph node and liver metastasis of PanNETs in RIP-Tag; RIP-tva mice.
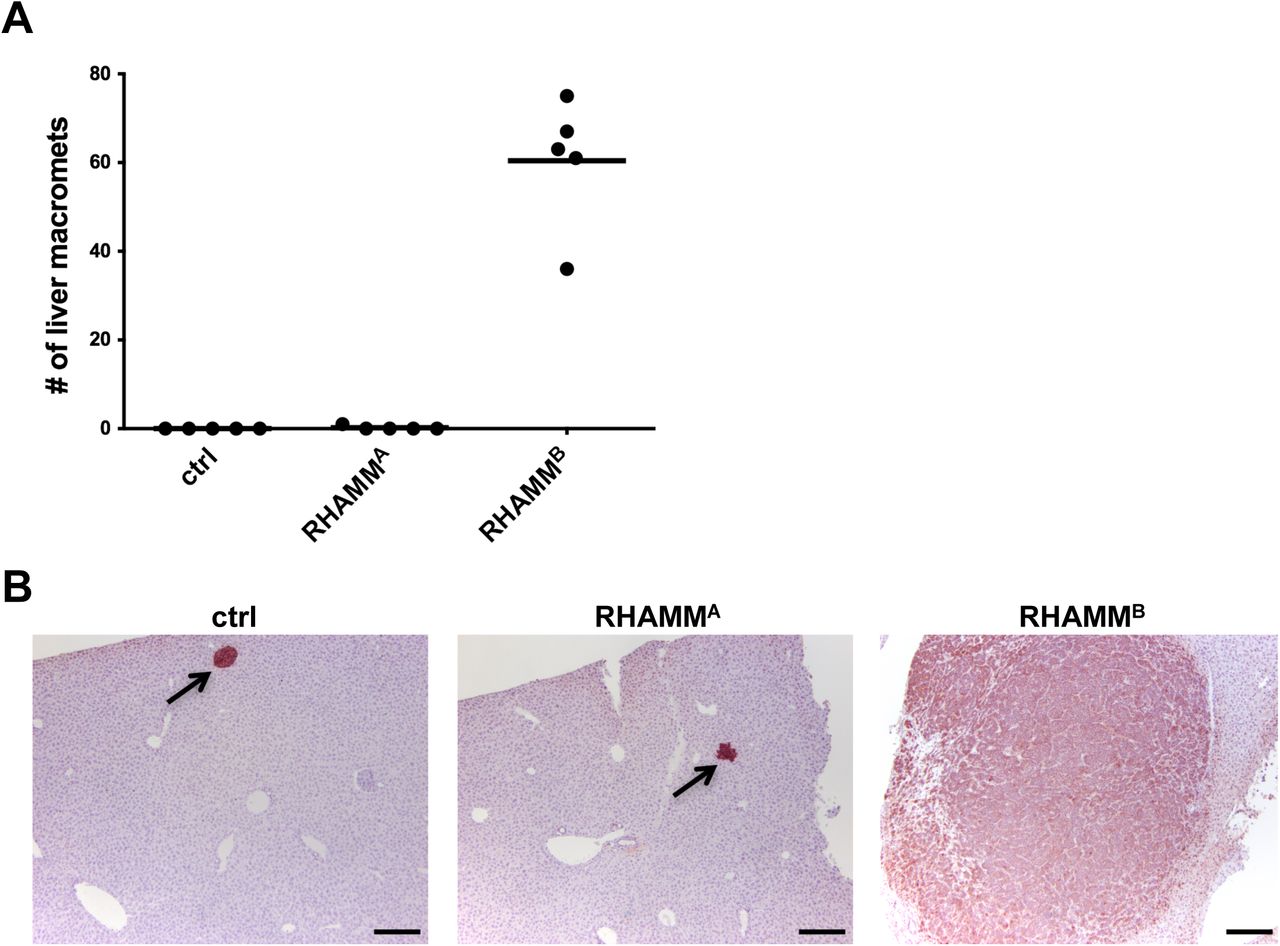
Unlike RHAMMB, RHAMMA did not promote liver metastasis of mouse PNET N134 cell line in a tail vein experimental metastasis assay. (A) The number of liver macrometastases was recorded. (B) Immunohistochemical staining of synaptophysin in the liver sections to reveal the presence of metastatic PNETs. Arrows indicate micrometastases. Original magnification: 10X. Scale bar, 200 µm.
RHAMMB is crucial for the metastatic potential of human PNET cell line BON1-TGL
BON1 is the most utilized human PNET cell line and was established from a peri-pancreatic lymph node in a patient with metastatic PNET. We found that BON1 had much higher expression of RHAMMB than RHAMMA as determined by RNA-Seq (Fig. 2A). We performed shRNA-mediated knockdown of total RHAMM to investigate whether this reduces metastasis of BON1-TGL cells, which carry the thymidine kinase/green fluorescent protein /luciferase fusion reporter (TGL). Knockdown of RHAMM by shRNA was confirmed (Fig. 2B-C). We used an orthotopic model of PNET liver metastases by injecting cells into the spleen of NSG mice [6]. Mice receiving control cells developed an average of 82.5 liver metastases after 3 weeks, while mice receiving BON1-TGL-shRHAMM cells developed an average of 17 liver metastases with significantly lowered tumor burden (Fig. 2D-F).

RHAMMB is crucial for metastatic potential of human PNET cell line, BON1-TGL. (A) RHAMMA and RHAMMB expression in BON1 cell line compared to those in normal islets. (B) RHAMMA and RHAMMB knockdown in BON1-TGL cell line by shRHAMM as determined by RT-qPCR analysis. (C) Western blot analysis of RHAMM and tubulin (as a loading control) in BON1-TGL-shLacZ cell line and BON1-TGL-shRHAMM cell line. (D-F) RHAMM knockdown greatly inhibited liver metastasis of BON1-TGL cells. A total of 0.5 million each BON1-TGL-shLacZ (control) or BON1-TGL-shRHAMM were injected into the spleen of NSG mice (n = 4 for each group). After 3 weeks, the recipient mice were euthanized to survey for metastatic sites and incidence. The number of liver macrometastases (D) and the tumor burden of liver macrometastases (E) were recorded. *: statistically significantly different (p < 0.05, one-tailed Mann-Whitney U test). Error bars in this figure represent standard deviation. (F) Liver sections with hematoxylin and eosin stain. Dashed circles indicate metastases. Original magnification: 10X. Scale bar, 1 mm. (G) RHAMMB overexpression in BON1-TGL-RHAMMB cell line. Western blot analysis of RHAMM and tubulin (as a loading control) are shown. (H) Representative bioluminescent images of NSG mice 4 weeks after injection (upper panel) and their organs (lower panel). A total of 1 million cells was injected into NSG mice via intracardiac injection (n = 7). (I) The tumor burden of macrometastases at adrenal glands was documented. *: statistically significantly different, p<0.05, t-test.
To determine whether even higher RHAMMB levels would further enhance metastasis of BON1-TGL cells, we generated BON1-TGL cells overexpressing RHAMMB (Fig. 2G). We injected cells into NSG mice via intracardiac injection. The increased levels of RHAMMB enhanced metastasis of BON1-TGL in mice throughout the mouse body as visualized by bioluminescence imaging and signals from multiple organs were higher in mice receiving BON1-TGL-RHAMMB than those in mice receiving BON1-TGL overexpressing a control vector (Fig. 2H). Notably, we observed macrometastases at the adrenal glands of mice receiving BON1-TGL-RHAMMB, but not in control mice (Fig. 2I). Taken together, RHAMMB is crucial for PNET metastasis.
RHAMMB is upregulated in human pancreatic ductal adenocarcinoma (PDAC) and correlates with poor survival
PDAC is the most common pancreatic cancer type. It was shown that total RHAMM is upregulated in primary PDAC by RT-qPCR of 14 matched tumors and adjacent normal tissues [7]. To compare RHAMMA and RHAMMB levels in PDAC, we analyzed publicly available The Cancer Genome Atlas (TCGA) datasets. Both RHAMMA and RHAMMB were expressed at significantly higher levels in PDAC than in normal pancreatic tissues, and RHAMMB was substantially higher than RHAMMA (Fig. 3A). Survival analysis showed that high RHAMM levels were correlated with a worse outcome (Fig. 3B). Furthermore, patients with high RHAMMB had inferior survival compared to those with high RHAMMA (Fig. 3C-D), suggesting that RHAMMB, but not RHAMMA, is a prognostic factor for survival of PDAC patients. Due to limited information available, we could not perform survival analysis for PNETs.


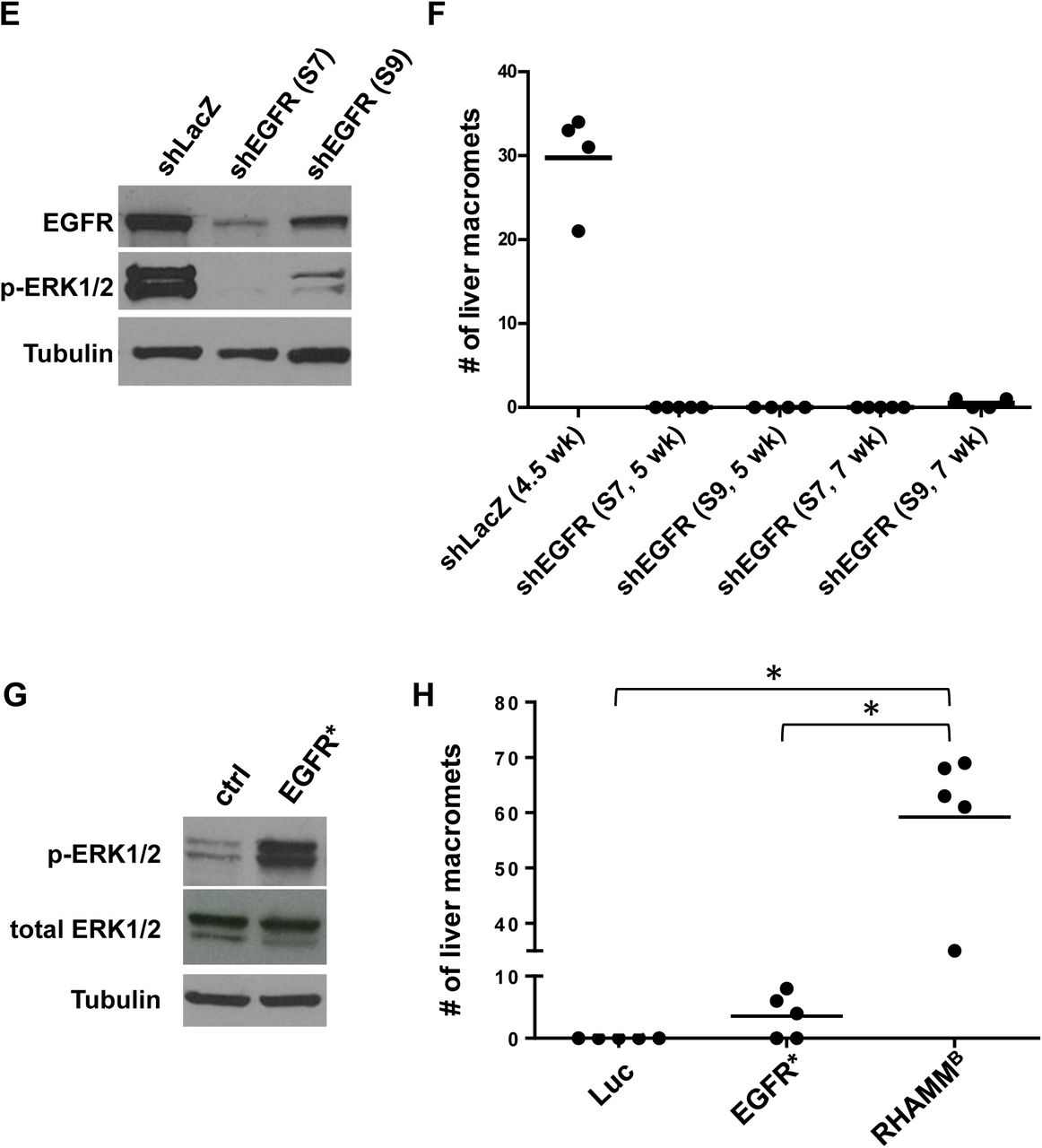
RHAMMB is upregulated in human pancreatic ductal adenocarcinoma (PDAC) and correlates with poor survival. (A) RHAMMA and RHAMMB expression values from TCGA PDAC dataset. RHAMMA: uc003lzf or NM_012484. RHAMMB: uc003lzg or NM_012485. cancer (ca): n = 124; control (ctrl): n = 4. Bars and error bars represent means and standard errors. (B-D) Kaplan-Meier survival analysis of TCGA cohort with 93 PDAC cases. High and low represent the status of the RHAMM mRNA expression levels compared to average values. (E) Knockdown efficiency of two EGFR shRNAs. S7 and S9 reduced EGFR protein expression and p-Erk1/2 levels in N134 cells overexpressing RHAMMB by Western blot analysis. α-tubulin was used a loading control. (F) EGFR knockdown greatly inhibited the liver metastasis of N134 cells overexpressing RHAMMB. A total of 1 million N134_RHAMMB_shLacZ, N134_RHAMMB_shEGFR(S7), or N134_RHAMMB_shEGFR(S9) cells were injected into the tail vein of NSG mice (n = 5 for each group). At the indicated time points, the recipient mice were euthanized to survey for metastatic sites and incidence. The number of liver macrometastases was recorded. (G) Western blot analysis of p-Erk1/2 and total Erk from N134 overexpressing luciferase (control), and N134_EGFR*. (H) N134 cells overexpressing luciferase (Luc), N134_EGFR* (EGFR*), N134_RHAMMB (RHAMMB) were injected into the tail vein of NSG mice (n = 5, each group). Five weeks later (for Luc and EGFR* groups) or when mice were lethargic (for RHAMMB), mice were euthanized to survey for metastatic sites and incidence. *: p< 0.0001, One-way ANOVA and pairwise comparison with Tukey’s adjustment.
EGFR signaling is required for RHAMMB-induced metastasis
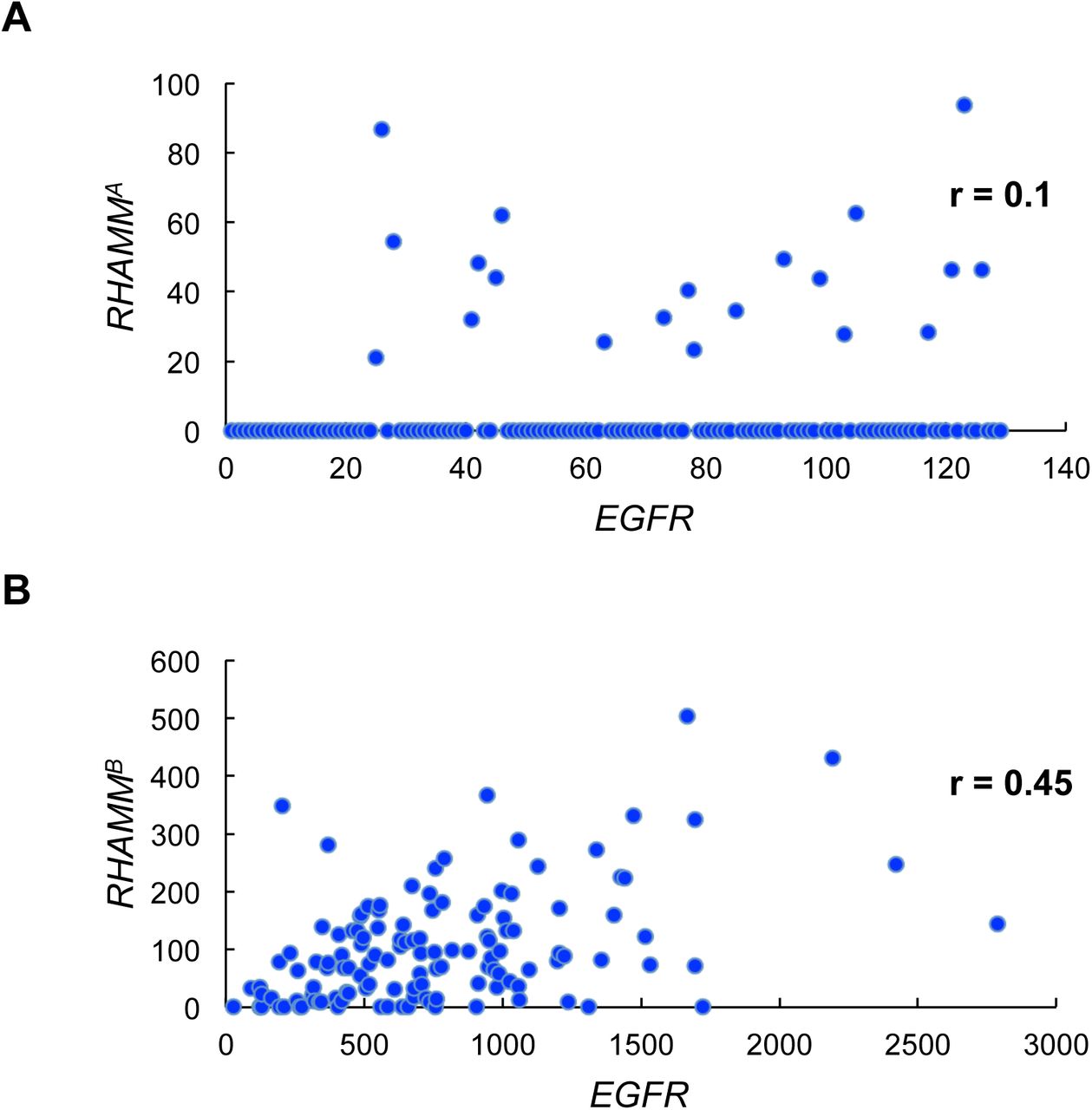
EGFR activation is associated with worse survival in many malignancies. Analysis of TCGA PDAC dataset showed a good correlation between EGFR and RHAMMB expression, but not between EGFR and RHAMMA expression (Fig. S3A-B). We previously showed that EGFR signaling is activated in N134-RHAMMB cells and an EGFR inhibitor, gefitinib, induces apoptosis of N134-RHAMMB cells [2]. These findings led us to hypothesize that enhanced EGFR signaling involves RHAMMB-induced metastasis. To test this hypothesis, we used two different shRNAs targeting mouse EGFR (S7 and S9) as well as a control shRNA targeting LacZ (shLacZ). shEGFR(S7 and S9) decreased the levels of both EGFR and p-ERK1/2, a downstream target of EGFR signaling, with S7 exhibiting a better knockdown efficiency than S9 (Fig. 3E).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlation between EGFR expression and RHAMMA (A) or RHAMMB (B) expression in TCGA PDAC dataset (RNA-Seq V2).
In tail vein metastasis assays, mice receiving N134-RHAMMB-shLacZ became lethargic, showing a mean of 29.8 liver macrometastases per mouse within 5 weeks, but mice injected with N134-RHAMMB-shEGFR(S7) and N134-RHAMMB-shEGFR(S9) cells did not develop metastases 5 weeks post-injection (Fig. 3F). An additional cohort of mice injected with N134-RHAMMB-shEGFR(S7) still did not develop metastases 7 weeks post-injection, and only a mean of 0.5 liver macrometastases appeared in mice injected with N134-RHAMMB-shEGFR(S9) at this time point (Fig. 3F).
We investigated whether a constitutively active form of EGFR, EGFR*, is sufficient to recapitulate RHAMMB activity in metastasis. Five of the 8 RIP-Tag; RIP-tva mice receiving RCASBP-EGFR* developed pancreatic lymph node metastases (62.5%) and 2 developed liver metastases (25%) (Table S1). We generated N134 cells overexpressing EGFR* (N134-EGFR*) for experimental metastasis (Fig. 3G). While no metastasis was found in mice receiving control N134-Luciferase after 6 weeks, an average of 6 liver macrometastases was detected in mice receiving N134-EGFR* (Fig. 3H). Although EGFR* promotes liver metastasis of PNETs in both spontaneous and experimental metastasis mouse models, the degree of metastasis is less than that of RHAMMB (Table S1 and Fig. 3H). Therefore, these EGFR knockdown and EGFR* overexpression data suggest that activation of EGFR contributes to RHAMMB-induced PNET metastasis, but EGFR* cannot fully recapitulate the metastatic phenotype of RHAMMB. Further studies are required to identify signals other than EGFR provided by RHAMMB and to understand whether endogenous levels of RHAMMA activate EGFR signaling.
Conclusion
We provide evidence that upregulation of RHAMMB is a valuable prognostic marker for pancreatic cancer. We demonstrated that only the shorter isoform RHAMMB, but not RHAMMA with 15 extra amino acids encoded by exon 4, is significantly upregulated in PNET and PDAC. RHAMMB, but not RHAMMA, promotes metastasis in spontaneous and experimental metastasis mouse models of PNET. EGFR signaling is required for RHAMMB-induced liver metastasis, but is not sufficient to promote PNET metastasis. RHAMMB, but not RHAMMA, is correlated with inferior survival in PDAC patients.
Methods and Materials
Bioinformatics and Statistical Analysis for RHAMM variants from RNA-Seq data
Total RNAs from 27 primary human PanNETs, 12 metastatic human PanNETs, and human BON1 cell line [8] were isolated with RNeasy Plus Universal Kits (Qiagen Cat no. 73404, Germantown, MD) according to the manufacturer’s protocol. Library preparation and RNA sequencing with paired-end 75 bp reads were performed according to protocols at the Genomics Resources Core Facility, Memorial Sloan Kettering Cancer Center, and Weill Cornell Medicine. A RNA-Seq dataset of 89 human pancreatic islets was obtained from publicly available database Gene Expression Omnibus (GEO accession GSE50398, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE50398) [9-11], and was converted to fastq file from SRA format using SRA Toolkit (v 2.4.4). All samples analyzed were aligned to hg19 reference genome using STAR (v 2.4) aligner. Aligned samples were then quantified to obtain gene expression in terms of FPKM (Fragments per Kilobase of Transcripts per Million Reads) against a UCSC hg19 annotation Gene Transfer Format containing coordinates for all genes, including specific RHAMM isoforms of interest using CuffLinks (v 2.2.1). FPKM values were extracted from isoform specific quantification output obtained from CuffLinks for each sample, with focus on isoform A (NM_012484) and isoform B of RHAMM (NM_012485). Log-transformed FPKM values (log2 [FPKM+1]) were used for further analysis to compare the expression levels across tissue types (Normal, Primary and Liver metastasis) and RHAMM isoforms (RHAMMA and RHAMMB) using two-way ANOVA, followed by pairwise comparisons with Tukey’s post-hoc test for multiple comparison adjustment. All analyses were performed in open-source data analysis software R (v 2.14.1) or SAS 9.4 (SAS Institute, Cary, NC).
Bioinformatics and statistical analyses were conducted on the publically available gene expression dataset from The Cancer Genome Atlas (TCGA; http://cancergenome.nih.gov/). TCGA normalized expression values were created by Illumina RNA-Seq version 2 RNA sequencing data (level 3). Downloaded data was analyzed using R or statistical software Prism (version 6.0f) for statistical computation and Kaplan-Meier survival analysis with log-rank test. Two-tailed Mann-Whitney U test was used to compare differences between two groups selected. P values < 0.05 were considered as statistically significant.
Quantitative real-time reverse transcription PCR (RT-qPCR)
Messenger RNA (mRNA) was isolated from human PanNET specimens or cells grown on 6-cm or 10-cm plates using RNeasy mini kit (Qiagen) containing gDNA eliminator spin columns. The cDNA was generated using SuperScript III First-strand synthesis system with random hexamers (Invitrogen), and power SYBR green (Invitrogen)-based qPCR was performed with 3 internal control genes and the comparative CT method (ΔΔCT).
The sequences of the primers used are RHAMMA (forward: located within exon 3, 5’-TGACAAAGATACTACCT TGCCTGCT-3’, reverse: located at the junction of exon 3 and 4, 5’-TCATTCTTTTGAGATTCCTTTGATTC-3’); RHAMMB (forward: located at the junction of exon 3 and 5, 5’-AAAGTTAAGTCTTCG GAATCAAAGATT-3’, reverse: located within exon 5, 5’-GCATTATTTGCA GAGAGAGATGT-3’), and internal control genes: human HMBS (forward: 5’-CCATCATCCT GGCAACAGCT-3’, reverse: 5’-GCATTCCTCAGGGTGCAGG-3’); human EEF1A (forward: 5’-CAATGTGGGCTTCAA TGTCAA-3’, reverse: 5’-CATAGCCGGCGCTTATTTG-3’); human MRPL19 (forward: 5’-GGGATTTGCATTCAGAGATCAGG-3’, reverse: 5’-CTCCTGGACCCGAGGATTATAA-3’).
Tissue preparation, immunohistochemistry, and scoring of protein expression
Retrospective and prospective review of PanNETs was performed using the pathology files and pancreatic cancer database at the authors’ institutions with Institutional Review Board (IRB) approval. Construction of TMA of human PanNETs was described previously [12]. Mouse tissues were fixed in 10% buffered formalin overnight at room temperature. Fixed tissues were processed and cut into 5 µm sections at Histoserv. Formalin-fixed/paraffin-embedded sections were deparaffinized and rehydrated by passage through a graded xylene/ethanol series before staining. Immunochemistry was examined by VECTASTAIN Elite ABC Kits (Vector Laboratories, Inc., Burlinggame, CA) following manufacturer’s instructions. The primary antibodies used were rabbit anti-RHAMM (1:1,000, Y800 [2], 1:100), rabbit anti-synaptophysin (1:100, Vector Laboratories, VP-S284 or 1:100, Lab Vision/neomarkers, Fremont, CA, RM-9111), and GFP (1:300, Invitrogen, Carlsbad, CA, A11122). RHAMM expression for each tumor was given a score of 0 if no staining was present, and a score of 1 if moderate to strong staining was present.
Cloning of RCASBP and retroviral vectors
RCASBP is a replication-competent avian leucosis virus with a splice acceptor and the Bryan-RSV pol gene. RCASBP-RHAMMB [2] and RCASBP-EGFR* [13] have been described. RCASBP-RHAMMA was generated using QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technology, Santa Clara, CA) to add exon 4 (forward primer: 5’-agttaagtcttcggaatcaaaggaatctcaaaagaatgataaa gatttgaagatattagagaaagagattcgtgttcttctacaggaac-3’, reverse primer: 5’-gttcctgtagaagaacacgaatctctttctctaatatcttcaaatctttatcattcttttgagattcctttgattccgaa gacttaact-3’) from RCASBP-RHAMMB. The presence of exon 4 in RCASBP-RHAMMA was confirmed by DNA sequencing.
The shRNA knockdown
Hairpin sequences targeting EGFR are S7: CCAAGCCAAATGGCATATTTA and S9: GCTTTCGAGAACCTAGAAATA. Hairpin sequence targeting LacZ is TCGTATTACAACGTCGTGACT. Hairpin sequence targeting RHAMM is sh1: GCCAACTCAAATCGGAAGTAT (Sigma, Clone ID: NM_012484.2-2128s21c1). Lentiviruses harboring shRNA were generated using 293T cells as described previously (http://www.broadinstitute.org/rnai/public/resources/protocols).
Cell culture and Western blot
Generation of N134 and BON1-TGL cell lines has been described [1, 14]. Human pancreatic neuroendocrine tumor cell line, BON1, was provided by Chris Harris [8, 12]. DF1, N134, and BON1-TGL were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 6 mM L-glutamine, and penicillin/streptomycin. The BON1-TGL cell line was transfected with pBABE-RHAMMB with lipofectamine 2000 (Invitrogen) to generate BON1-TGL-RHAMMB cell line. BON1-TGL cells overexpressing a control vector or RHAMMB were cultured in DMEM supplemented with 0.5 µg/ml puromycin, 10% FBS, 6 mM L-glutamine, and penicillin/streptomycin. N134 cells overexpressing cDNAs were generated as described [15]. N134 cells overexpressing shRNAand BON1-TGL overexpressing shRNA were cultured in medium supplemented with 0.4 µg/ml puromycin and 0.5 µg/ml puromycin, respectively.
For Western blot analysis, cell extracts were loaded into mini-protean 4-15% pre-cast gels (Bio-Rad). Protein was immobilized onto nitrocellulose membrane, 0.45 µm pore size (Bio-Rad). Blots were blocked for one hour with 3% (weight/volume) bovine serum albumin (Fisher Scientific) and incubated overnight at 4° C with a rabbit monoclonal antibody to RHAMM [EPR4055] antibody at 1:1,000 dilution (Abcam, ab108339). The next day blots were washed 4 times for 10 minutes with TBST and incubated for one hour at room temperature with an anti-rabbit secondary antibody at 1:5,000 dilution. Bands were detected with enhanced chemiluminescence.
Animal experiments
Generation of RIP-Tag; RIP-tva mice has been described [1] and detailed protocols for somatic delivery of the RCASBP viruses has been described [15]. Immunodeficient mice, NOD/scid-IL2Rgc knockout (NSG), were generated by the Jackson Laboratory. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All mice were housed in accordance with institutional guidelines. All procedures involving mice were approved by the Institutional Animal Care and Use Committee of Weill Cornell Medicine. For the experimental metastasis assay, either 1 x 106 N134 cells in 100 µL PBS were injected into the tail veins of NSG mice or 1 x 106 BON1-TGL cells in 100 µL PBS were injected into the left ventricle of NSG mice. The orthotopic model of PanNET liver metastasis was performed as previously described [16] with the following modification: 0.5 x 106 BON1-TGL cells in 50 µL DMEM containing 2% FBS were injected into the spleen. A standard formula for tumor volume was applied (volume [mm3] = 0.52 × width2 × length). Tumor burden is the sum of the tumor volume per mouse.
Abbreviations
- RHAMM
- receptor for hyaluronic acid-mediated motility
- PNET
- pancreatic neuroendocrine tumor
- TCGA
- The Cancer Genome Atlas (TCGA)
- HA
- hyaluronic acid
- FPKM
- fragments per kilobase of transcripts per million reads
- RT-qPCR
- quantitative real-time reverse transcription PCR
- PDAC
- pancreatic ductal adenocarcinoma
- DMEM
- Dulbecco’s modified Eagle’s medium
- FBS
- fetal bovine serum.
Declarations
Ethics approval and consent to participate
All procedures involving mice were approved by the institutional animal care and use committee. Retrospective and prospective review of PNETs was performed using the pathology files and pancreatic cancer database at the authors’ institutions with IRB approval.
Consent for publication
All authors agreed on the manuscript.
Availability of data and material
The RNA-Seq and TCGA datasets that support the findings of this study are available in Gene Expression Omnibus (GEO accession GSE50398, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE50398) [9-11], and http://cancergenome.nih.gov/cancersselected, and the other data are available form the authors upon reasonable request.
Competing interests
The authors declare no conflict of interest.
Funding
This work is partially supported by NIH grants 2U01DK072473 (to Y.-C.N.D.), 1R21CA173348-01A1 (to Y.-C.N.D.), 1 R01CA204916-01A1 (to Z.C., G.Z., Y.-C.N.D.), UL1TR000457 (to Z.C.), DOD grants W81XWH-13-1-0331 (to S.C., Z.C., Y.-C.N.D.) W81XWH-16-1-0619 (to Y.-C.N.D.), and Goldhirsh Foundation (to A.V., T.J. F.III, O.E., Y.-C.N.D.).
Authors’ contributions
S.C., X.C., B.J.K., G.Z., S.P., T.H., and L.S. performed the experiments and analyzed the data. D.W. conducted the analysis on the TCGA dataset. D.W., G.Z., S.P., T.J. F.III, and K.R. edited the manuscript. L.H.T. contributed immunohistochemical interpretation. A.V., C.S.C., and O.E. contributed to bioinformatics analyses. Z.C. performed statistical analysis. L.H.T., K.R., and T.J. F.III contributed to samples. Y.-C.N.D. designed and performed the experiments, analyzed the data, and wrote the manuscript.
Acknowledgments
The authors thank Danny Huang for mouse database design; Harold Varmus, Yi Li, Jihye Paik, Todd R. Evans, David Foster, Bi-Sen Ding, Katie Politi, Jeffrey Yongchun Zhao, Romel Somwar, Selina Chen-Kiang, Stephanie Azzopardi, Samantha Li, Megan Wong, Joseph Na, and Robin Zhang for their valuable input and excellent assistance; Diane L. Reidy and David S. Klimstra for contributing the approval of the tumor materials.
References




