

Abstract
Cyanobacteria produced the atmospheric O2 that began accumulating 2.4 billion years ago1, leading to Earth’s Great Oxidation Event (GOE)2. For nearly 2 billion years following the GOE, O2 production was restricted and atmospheric oxygen remained low2–5. Oxygen rose again sharply with the advent of land plants roughly 450 million years ago, which increased atmospheric O2 through carbon burial4–5. Why did the O2 content of the atmosphere remain constant and low for more than a billion years despite the existence of O2-producing cyanobacteria? While geological limitations have been explored2–7, the limiting factor may have been biological, and enzymatic. Here we propose that O2 was kept low by oxygen inhibition of nitrogenase activity. Nitrogenase is the sole N2-fixing enzyme on Earth, and is inactive in air containing 2% or more O2 by volume8. No O2-resistant nitrogenase enzyme is known9–12. We further propose that nitrogenase inhibition by O2 kept atmospheric O2 low until upright terrestrial plants physically separated O2 production in aerial photosynthetic tissues from N2 fixation in soil, liberating nitrogenase from inhibition by atmospheric O2.
Current views of oxygen in Earth history (Fig. 1) depict the first traces of O2 appearing in the atmosphere starting about 2.7 to 2.5 Gy ago1–5. During the Great Oxidation Event, or GOE, roughly 2.4 billion years ago2, O2 rose to about 10% of its present atmospheric level (PAL), corresponding to an atmosphere of roughly 2% O2 by volume2, or even less3. Isotopic studies indicate that for roughly 1.5 billion years following the comparatively sudden GOE, further net O2 accumulation ceased, with atmospheric levels remaining stable and below 10% PAL2–4.

Modified after refs. 1-5. For most of the Proterozoic eon, free O2 was much less abundant than it is today. Lyons et al. (2014) estimate Proterozoic O2 in the atmosphere as low as 0.001 PAL while Holland2 estimates atmospheric O2 at around 0.1 to 0.2 PAL. Stolper and Keller4 estimate mid-Proterozoic deep ocean dissolved O2 concentrations at about 11 µM or roughly 0.06 of the present value of 178 µM. “Whiffs” refers to isotope signatures for evidence of transient, local O2 before the GOE1,3. The Lomagundi excursion is represented as a dotted line because it is included in the summary of ref.3 but not in that of refs.1,2,4,5. Heterocysts are differentiated cells of some cyanobacteria, and protect nitrogenase from inactivation by O2. Their relevance is that cyanobacteria have an ancient fossil record, but the oldest fossil heterocysts26 are younger than land plants, suggesting that cyanobacteria evolved this mechanism of O2 protection in response to Phanerozoic O2 accumulation. PAL; Present Atmospheric Level. GOE; Great Oxidation Event.
With atmospheric O2 low, marine O2 stayed low as well. Geochemical evidence suggests that the oceans remained largely anoxic throughout the Proterozoic1–5, with a rapid rise to roughly modern oxygen levels starting around 580 My ago, perhaps as recently as only 430 My ago4,5 (Fig. 1). Late increases in atmospheric O2 implicate the emergence of land plants and terrestrial carbon burial as a causal factor. Today, land plants comprise roughly 97% of Earth's surface-exposed biomass7. Their ecological success has been linked to the rise in O2 because terrestrial sequestration of organic carbon as fibrous biomass protects it from reoxidation to CO2, curbing O2 consumption4–5.
What limited oxygen accumulation? A major puzzle of O2 history is why O2 rose so late, that is, why atmospheric and marine O2 levels stayed low for almost 2 billion years despite the existence of cyanobacteria, which were capable of continuous light-driven O2 production. What held cyanobacteria back, why did O2 stop accumulating after the GOE and why did it remain low during the Proterozoic, or the “Boring Billion” as it is sometimes called3.
Geochemists have long recognized that Proterozoic O2 stasis presents a problem and have proposed a number of explanations to account for the delayed oxygen rise. Some proposals posit a steady supply of geochemical reductants from within the Earth, such as Fe2+ or S2–, reductants that continuously consumed the O2 produced by cyanobacteria, keeping O2 low8. Other proposals invoke biotically induced changes affecting in the degree of mixing between nutrient rich reservoirs and the photic zone, for example through animal burrowing activity9. Germane to many proposals is the concept that crucial nutrients such as molybdenum, which is required for nitrogenase activity, were limited in supply by geochemical factors and that nitrogenase limited primary production for this reason6. These proposals and others2–5 might apply to some areas of the ocean or some phases of Earth’s history. But how and why any set of factors should act in concert to keep O2 low for almost 2 billion years is yet unresolved10.
Nitrogenase regulates oxygen levels. We propose that O2-dependent feedback inhibition of a single enzymatic activity limited O2 accumulation during the boring billion: inhibition of nitrogenase by O2 gas. Carbon and nitrogen enter the biosphere in distinct chemical reactions catalysed by specific enzymes. For carbon there are six pathways of CO2 assimilation that differ in age, oxygen tolerance, and key CO2 reducing enzymes11. For N2 there is only one entry point into metabolism: nitrogenase12–14. Nitrogenase is widespread among cyanobacteria12. There are Mo, Fe and V containing isoforms of the enzyme that all share a common ancestor and homologous active sites13–15.
The nitrogenase active site is replete with metal cofactors (Fig. 2) and harbours a metal coordinated carbide carbon atom, unique among all enzymes known so far14. Like a blacksmith, nitrogenase uses ancient but robust technology. Nitrognease has an obligatory H2 producing side reaction, and it requires 8 electrons and 16 ATP per N2 fixed, the ATP being consumed at steps that alter the redox potential of FeS clusters via conformational change13. Nitrogenase requires numerous assembly factors14, and has been neither replaced nor improved during evolution, which reveals that the solution that life found to fix N2 is the only one readily attainable in 4 billion years of physiological engineering by microorganisms. Nitrogenase is a limiting factor. It is inhibited by O2 in a feedback loop (Fig. 3), and this simple property alone could limit O2 accumulation over geological time.

Redrawn from ref.15 with the proposed binding site for N2.
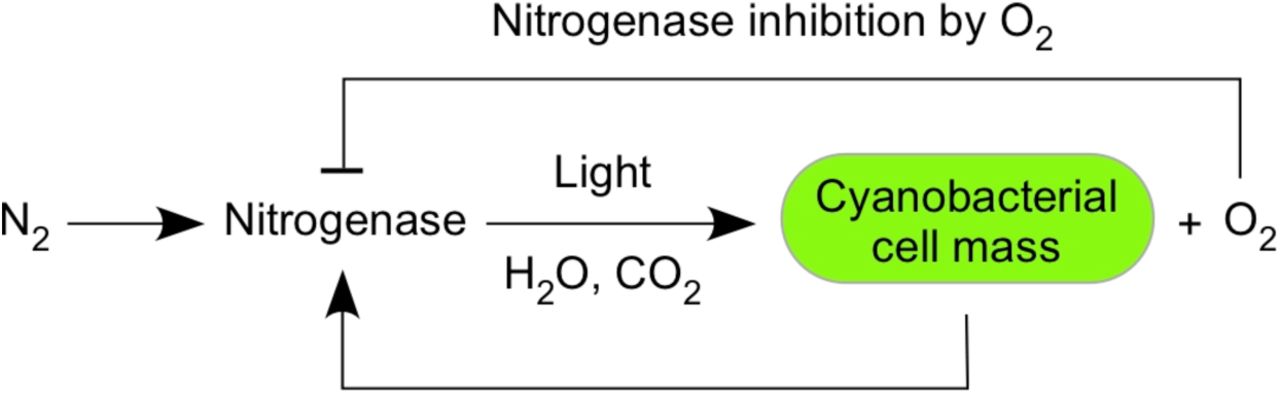
O2 inhibits nitrogenase, which is required for O2 production in photosynthesis. A steady state is reached at environmental O2 levels not exceeding 10% PAL.
Nitrogenase feedback inhibition operates as follows. By dry weight, cells are about 50% carbon and about 10% nitrogen. Cyanobacteria had water as an unlimited reductant for CO2fixation, but, for net growth to occur, N2 incorporation had to keep pace. Nitrogenase is inhibited by oxygen, the product of water oxidation — but there is a threshold of oxygen concentration below which nitrogenase remains active and above which nitrogen fixation ceases completely. If diazotrophic cyanobacteria are grown under conditions where they have sufficient CO2 and light, and with N2 as the sole N source, then they grow and accumulate no more than 2% oxygen in their culture atmosphere16. The 2% O2 remains constant during prolonged culture growth because this is the O2 partial pressure beyond which nitrogenase activity becomes inhibited. With greater O2, nitrogenase is inactivated and there is no fixed N to support further biomass accumulation. With less O2, nitrogenase outpaces CO2 fixation until the latter catches up, returning O2 to 2% in the culture. In cyanobacteria, CO2 fixation means O2 production.
In microbial mats, oxygen inhibits nitrogenase activity and nitrogenase gene transcription following the onset of illumination during the natural diel cycle17. The initial effect of illumination, however, is to increase nitrogenase activity to its maximum value by means of increased ATP and reductant from photosynthetic electron transport. After a lag of a few hours, O2 concentration becomes inhibitory to nitrogen fixation17. Because there is no biochemical alternative to nitrogenase for fixing N2, because there are no O2 tolerant nitrogenases known, and because reductant for CO2 fixation was not limiting for cyanobacteria, this feedback loop would have operated, on a planetary scale, for two billion years or more. While primary production using H2S instead of H2O as in Oscillatoria limnetica18 is also subject to O2-feedback inhibition, it would have been limited via the availability of reductant at O2 levels far below those created by of oxygenic photosynthesis 19, and would not have impacted O2 accumulation. Nitrogenase is an O2 inhibited sensor that kept environmental O2 low throughout the Proterozoic.
Cyanobacteria have evolved mechanisms to avoid nitrogenase inhibition by oxygen12, including N2 fixation in the dark20, heterocysts21 or filament bundles as in Trichodesmium22. Critics might counter that any one of those mechanisms could have bypassed O2 feedback inhibition. There are three problems with this objection. First, evolution operates without foresight. Second, the mechanisms that cyanobacteria use to deal with modern O2 levels appear to have arisen independently in diverse phylogenetic lineages, not at the base of cyanobacterial evolution when water oxidation had just been discovered23,24. Third, the oldest uncontroversial fossil heterocysts trace to land ecosystems of the Rhynie chert and are merely Devonian in age25 (Fig. 1), suggesting that heterocysts arose late in evolution, probably in response to levels of O2 exceeding 2% by volume. Fossil akinetes — cyanobacterial resting spores — have been found in older sediments26,27, yet there is no direct evidence for heterocysts older than the first land plants.
The concept of limiting metal availability (Mo, V, or Fe) for nitrogenase activity6 is an element of many proposals to account for low Proterozoic O2. Our proposal differs from nutrient limitation in a crucial aspect. Limiting the number of active nitrogenase enzymes in the environment by limiting metal (Mo, V, and Fe) availability only limits the rate at which cyanobacteria produce O2, requiring other factors to impose limits upon the final O2 partial pressure. Nitrogenase feedback inhibition regulates the O2 partial pressure directly, independently of the rate of photosynthesis, and generates a value that corresponds to the geochemical observation.
An O2 overshoot 2.3 billion years ago is suggested by an isotopic anomaly called the Lomagundi excursion. At 2.3 to 2.2 Ga ago, the isotopic record first reported from the Lomagundi formation in Zimbabwe indicates burial of heavy (13C enriched) carbon3. This 13C increase is interpreted3, though not universally2, as indicating the presence of large amounts of O2 on a global scale. If that interpretation is correct, its least explicable aspect is that following the Lomagundi excursion, oxygen levels drop once again3. Yet they do not drop to pre-cyanobacterial levels, rather they drop to oxygen levels very near 2% O2, the oxygen partial pressure that nitrogenase feedback inhibition generates. If the Lomagundi excursion is taken as a valid proxy for high global O2 levels, the following situation prevailed at the GOE. O2 is a strong oxidant. Its contribution to metabolic evolution was not just new metabolic pathways, but more complete oxidation of existing organic substrates28. O2 mobilized organic nitrogen and carbon that had been sequestered in biomass. By liberating sequestered nitrogen (and carbon as CO2) that had previously been inaccessible to anaerobes, the onset of O2 accumulation at the GOE initiated a positive growth feedback loop for aerobic autotrophs that were not reductant limited: cyanobacteria. When anaerobically deposited nitrogen reserves had been liberated, nitrogenase feedback inhibition set in, driving O2 levels down to Proterozoic levels, and keeping them low for a billion years thereafter. Our proposal does not hinge upon the Lomagundi excursion, yet if the excursion is interpreted as evidence for transiently high global O2 levels then our proposal can account both for its emergence(nitrogenase independent N availability during the excursion) and for its decline in the subsequent return to low O2.
When and why did feedback inhibition at nitrogenase cease to keep O2 low? At the origin of land plants, the nature of biomass changed and O2 production by upright terrestrial plants became physically separated from N2 fixation in aquatic environments and soil. Deposition by land plants of nitrogen-depleted cellulose, billions of tonnes of it, became a massive sink for CO2 without exerting similar effects on nitrogen availability, thus allowing O2 to increase through the standard mechanism of carbon burial, bypassing control by aquatic nitrogenase feedback.
Critics might interject that O2 levels began to rise before the first fossil occurrence of land plants29. We point out that nitrogenase limitation determines the maximum O2 partial pressure near the water surface for nitrogenase-limited oxygen production. This limit does not identify the timepoint at which the deep ocean becomes fully oxic, since that depends upon other factors such as reductant load, ocean mixing, or both, independently of photic zone nitrogenase limitation. Stolper and Keller4 report that deep ocean oxygenation became complete 540 million years ago. If so, that was the first time (or possibly the first time since the Lomagundi excursion) that N-rich organic ocean floor sediment came into widespread contact with oxygenated water. This contact released organic N, leading to atmospheric O2 increase, after which O2 levels dropped once again29,30 to the value imposed by the nitrogenase limit. Nitrogenase inhibition returns O2 to low levels following O2 increases, thus explaining an otherwise puzzling aspect of Proterozoic O2 variation.
In conclusion, oxygen inhibition of any ecosystem’s cornerstone enzyme activity, nitrogenase, created a bottleneck for oxygenic primary production that is sufficient to account for low oxygen levels throughout the boring billion. Nitrogenase feedback inhibition could directly account for Proterozoic low oxygen stasis. It would have driven down transiently higher O2 levels ensuing from nitrogenase-independent N availability, and it would have ceased at the origin of land plants. Our model requires light, CO2 and N2 in the photic zone and hence accommodates local and global variation in geochemical conditions while remaining robust to their effects. We propose that the factor limiting Proterozoic O2 accumulation was not geochemical. It was biological, and the attribute of a single enzyme, nitrogenase, contained within and synthesized by living cells.
Acknowledgements
We thank Olivia P. Judson and Dan Wang for comments on the manuscript. JFA thanks the Leverhulme Trust for Research Fellowship EM-2015-068. WFM thanks the ERC (grant no. 666053), the Volkswagen Foundation (grant no. 93 046) and the DFG (grant no. 1426/21-1) for funding.
Footnotes
↵* e-mail: j.f.allen{at}ucl.ac.uk





{kind=link}
{kind=link}
{kind=link}