

Abstract
Cell differentiation and cell fate determination in sensory systems are essential for stimulus discrimination and coding of environmental stimuli. Color vision is based on the differential color sensitivity of retinal photoreceptors, however the developmental programs that control photoreceptor cell differentiation and specify color sensitivity are poorly understood. In Drosophila melanogaster, there is evidence that the color sensitivity of different photoreceptors in the compound eye is regulated by inductive signals between cells, but the exact nature of these signals and how they are propagated remains unknown. We conducted a genetic screen to identify additional regulators of this process and identified a novel mutation in the hibris gene. hibris encodes an irre cell recognition module protein (IRM). These immunoglobulin super family cell adhesion molecules include human neph and nephrin (NPHS1). hibris is expressed dynamically in the developing Drosophila melanogaster eye and loss-of-function mutations give rise to a diverse range of mutant phenotypes including disruption of the specification of R8 photoreceptors cell diversity. The specification of blue or green sensitivity in R8 cells is also dependent upon Notch signaling. We demonstrate that hibris is required within the retina, non-cell autonomously for these effects, suggesting an additional layer of complexity in the signaling process that produces paired expression of opsin genes in adjacent R7 and R8 photoreceptor cells.
Author Summary As humans, our ability to distinguish different colors is dependent upon the presence of three different types of cone cell neurons in the retina of the eye. The cone cells express blue, green or red absorbing visual pigments that detect and discriminate between these colors. The principle of color discrimination by neurons “tuned” to different colors is an evolutionarily conserved specialization that occurs in many different animals. This specialization requires 1) visual pigments that detect different colors and 2) a developmental program that regulates the expression of these pigments in different types of cells. In this study we discovered that the fruit fly (Drosophila melanogaster) gene hibris is required for the developmental program that produces blue sensitive neurons in the fly retina. When we over-expressed hibris throughout the developing retina, extra blue sensitive cells were produced. These results demonstrate that if there is not enough hibris, too few blue sensitive cells form, but if there is too much hibris, too many blue sensitive cells form. Finally, we discovered that the hibris gene does not act in color sensitive neurons of the retina themselves. This surprising discovery suggests that hibris may influence development of the retina in a completely new and different way.
Introduction
Color vision in humans and most other organisms is dependent upon the expression of spectrally distinct visual pigments (opsins) in different photoreceptor cells [1-3]. The organization of photoreceptor cells within the retinal mosaic reflects a variety of different developmental mechanisms, including regional specialization, stochastic, and precise cell-cell adjacency [4]. D. melanogaster is capable of color vision and is a useful experimental system for examining the developmental programs that produce photoreceptor cells having different color sensitivities [5-12]. The compound eye consists of ∼800 ommatidia, each containing eight rhabdomeric photoreceptor cells (R cells). The central R7 and R8 photoreceptor cells mediate polarization sensitivity and color vision [13, 14]. As shown in Fig 1, the majority of ommatidia contain matched pairs of R7 and R8 cells expressing specific rhodopsin (Rh) visual pigments, either Rhodopsin 3 (Rh3, FBgn0003249) and Rhodopsin 5 (Rh5, FBgn0014019) (tandem magenta-blue cylinders), or Rhodopsin 4 (Rh4, FBgn0003250) and Rhodopsin 6 (Rh6, FBgn0019940) (tandem yellow-green cylinders).
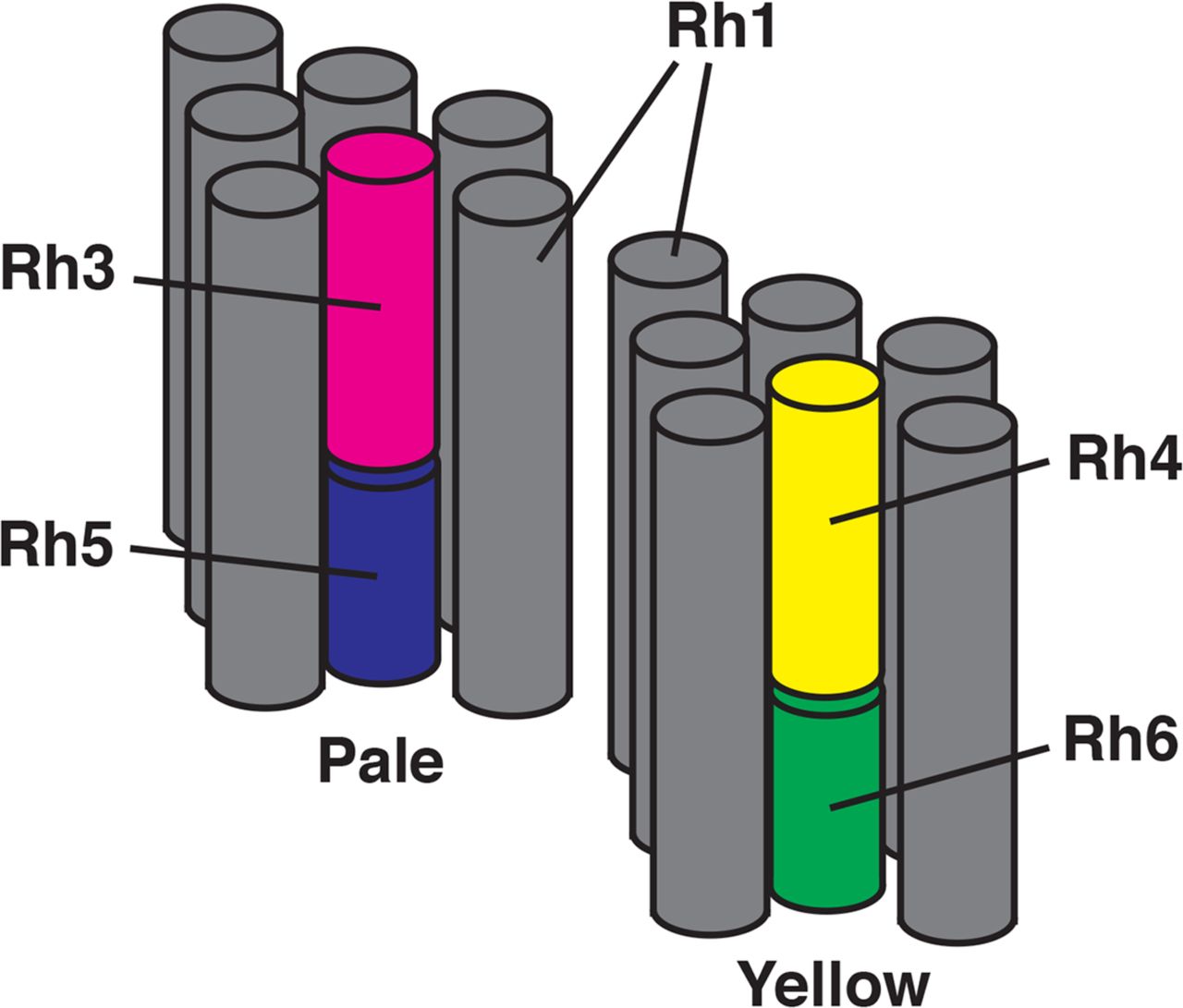
Two ommatidia are shown consisting of gray cylinders corresponding to the rhabdomeres of the R1-6 photoreceptor cells. These surround the central rhabdomeres of the R7 and R8 cells. Expression of opsin genes within the R7 cells (Rh3 in magenta or Rh4 in yellow) is paired with opsin gene expression in the adjacent R8 cell (Rh5 in blue or Rh6 in green) in pale and yellow ommatidia, respectively.
These two main ommatidial subtypes were initially identified based on pale or yellow fluorescence when illuminated with blue light [15, 16], with pale (pR7/pR8) expressing Rh3/Rh5, while yellow (yR7/yR8) cell pairs express Rh4/Rh6 (Fig 1) [10, 11, 17]. This paired expression of opsin genes in adjacent R7 and R8 cells within an individual ommatidium is thought to result from a series of developmental steps. First, a subset of R7 cells stochastically and cell autonomously express spineless (ss, FBgn0003513) which represses Rh3 and induces Rh4 expression [18]. In pR7 cells that stochastically fail to express ss and do express Rh3, a signal is initiated that induces the expression of Rh5 in adjacent pR8 cells. Extensive studies have identified the genes warts (wts, FBgn0011739), melted (melt, FBgn0023001), members of the hippo (hpo, FBgn0261456) pathway, along with the TGFβ superfamily receptors baboon (babo, FBgn0011300) and thick vein (tkv, FBgn0003726), and their respective ligands as components of the inductive signal from pR7 that drives the expression of Rh5 in pR8 [12, 19-21]. In the absence of a signal from yR7, the default yR8 fate and expression of Rh6 occurs. In addition, we have found that the Epidermal growth factor receptor (Egfr, FBgn0003731) and rhomboid (rho, FBgn0004635) are also required for this process [22, 23].
Here we undertook a genetic screen to identify additional genes required for this process and show that hibris (hbs, FBgn0029082), an irre Cell Recognition Molecule (IRM) [24], NPHS1 (nephrin, Homo sapiens, HGNC:9801) related member of the Immunoglobulin Super Family (IgSF), as well as Notch (N, FBgn0004647) are required for the establishment of paired opsin expression in adjacent R7 and R8 photoreceptor cells. Interestingly, we found that hbs is required non-cell autonomously for this process, suggesting the involvement of additional interactions between R7, R8 and neighboring cells.
Results
Isolation and characterization of the a69 mutant
To identify genes required for the induction of Rh5 expression in R8 photoreceptors, we screened a collection of approximately 150 homozygous viable eye-expressing enhancer trap lines carrying insertions of the P{etau-lacZ} transposon (FBtp0001352) [25]. This was based on the rationale that genes required for the induction of Rh5 expression would be expressed in the eye, the P{etau-lacZ} transposon has been especially useful in studies of the nervous system, and insertion of this element into loci of interest would provide a convenient means to identify the affected genes [25]. The percentage of Rh5-expressing R8 cells was determined by labeling dissociated ommatidia with antibodies against Rh5 and Rh6. Several mutants with abnormal percentages of Rh5-expressing R8 cells were noted and a69 (FBgn0026612), with the lowest percentage of Rh5 (9%) was further characterized. Immunostaining of both dissociated ommatidia and tissue sections showed that in the a69 enhancer-trap line, Rh5- expressing R8 cells are reduced and most R8 cells have assumed the default fate and express Rh6 (Fig 2A-E, Table 1). Since mutants lacking R7 cells or having a reduced number of Rh3 expressing R7 cells would also show diminished Rh5 expression, we examined the expression of the opsins expressed in the R7 cells and found that the percentage of Rh3 expressing R7 cells was similar to white1118 (w1118, RRID:BDSC_3605) control flies (41.9%, Table 1).However, there was a dramatic mispairing between Rh3 expressing R7 cells adjacent to Rh6 expressing R8 cells (Fig 2C,F, Table 1) compared to cinnabar1 brown1 controls (cn1 bw1, RRID:BDSC_264) consistent with the idea that the a69 enhancer trap line carries a mutation in a gene required for the induction of Rh5 expression in R8 cells.

White eyed wild type flies (w1118) express Rh5 and Rh6 in a proportion of approximately 1:2, this is shown in a longitudinal section of the retina (A) as well as in dissociated ommatidia (B). (C) w1118 flies express Rh4 and Rh6 in a paired fashion. The arrowheads indicate Rh3 expressing R7 cells that do not normally pair with Rh6 expressing R8 cells. w1118; P{etau-lacZ}a69 mutants show a disruption in Rh5 expression, with a substantial decrease in Rh5 expression shown in both section (D) and dissociated ommatidia (E) as well as prominent mispairing between Rh3 expressing R7 cells and Rh6 expressing R8 cells in the same ommatidia (arrowheads).
To isolate the gene responsible for the a69 phenotype, the location of the P-element insertion in a69 was determined and found to map to the right arm of the second chromosome at position 60E (data not shown). To determine whether the P-element in a69 is the cause of the phenotype, P-element excision lines were generated and analyzed. Thirty-five homozygous strains of these excision chromosomes were analyzed by staining dissociated ommatidia with antibodies against Rh5 and Rh6, and all of them (100%) were found to have a low Rh5 percentage, similar to that of a69 (data not shown). Only 1% of excision strains would be expected to retain the mutant phenotype as a result of imprecise excision, thus our inability to revert the mutant phenotype is consistent with the a69 P-element not being responsible for the mutation [26]. Furthermore, mapping via recombination analysis revealed that the a69 mutation is localized to the interval between the purple (pr, FBgn0003141) and curved (c, FBgn0000245) genes in the middle the second chromosome (Fig 3, S 1 Table), far away from the P-element insertion site in a69. From this we conclude that the a69 mutation is not associated with the insertion of the P-element. Thirty-three deficiency lines located in the region between pr and c were tested for a69 complementation (Fig 4, S 2 Table). These analyses narrowed the location of the a69 mutation to 51C3-51D1 (Fig 4). The lower portion of Fig 4 shows a diagram of this genomic region, spanning ∼300 Kb and encompassing 25 known protein coding genes.
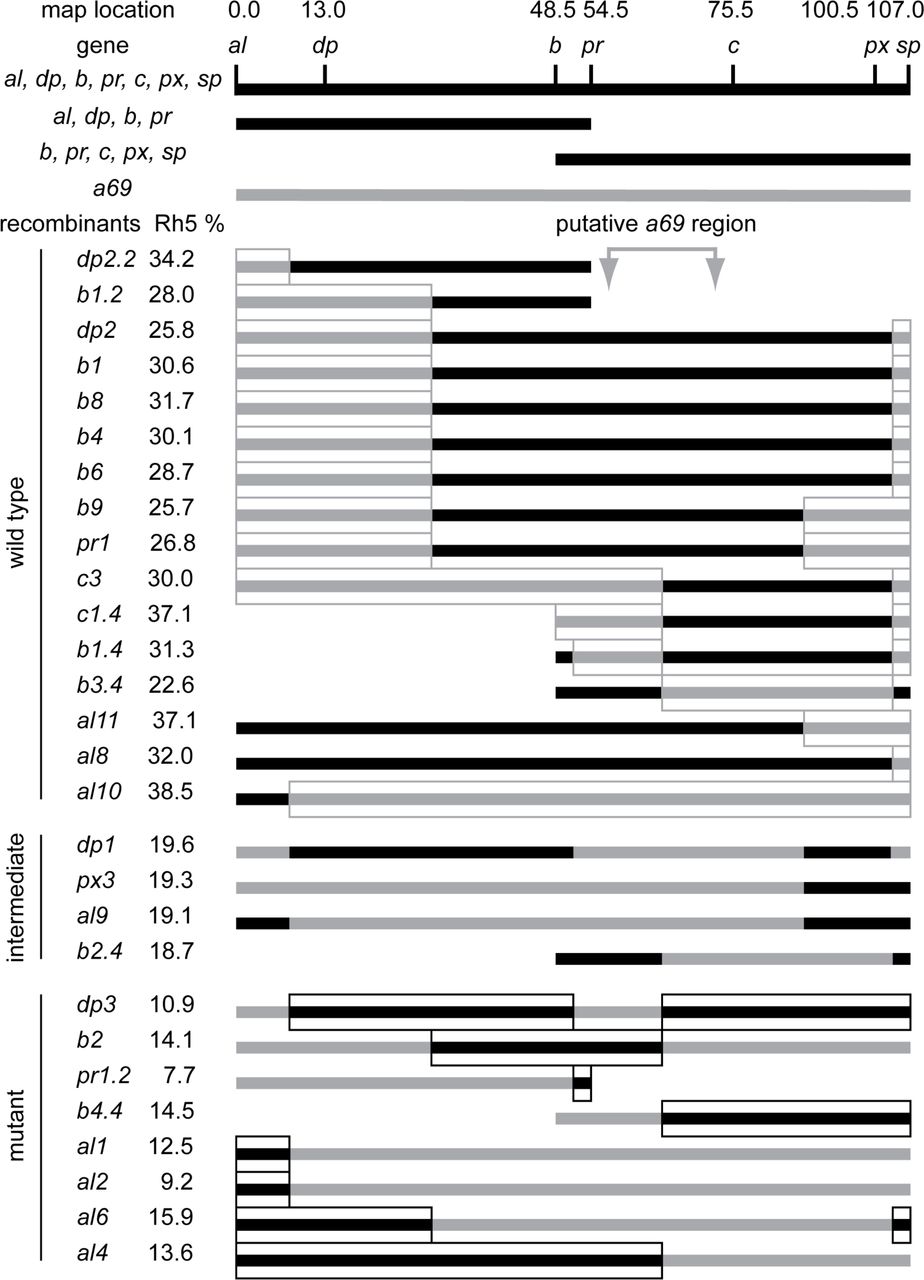
Three multiply marked chromosomes (al1 dpyov1 b1 pr1 c1 px1 sp1, al1 dpyov1 b1 pr1, and b1 pr1 c1 px1 sp1) were recombined with the w1118; P{etau-lacZ}a69 mutant. After marker identification, recombinant strains were back crossed to the a69 mutant and scored for the percentage of Rh5 expression. The regions of the recombinant chromosomes assumed to be derived from the a69 parental mutant strain are indicated in gray, while the regions assumed to be derived from the multiple marked (wild-type) chromosomes are black. Sixteen recombinant strains were phenotypically wild-type and complemented a69. Four recombinant strains were intermediate and eight strains were mutant and failed to complement a69. The four intermediate strains and one wild type strain, al10, differed from the expected phenotypes and may have resulted from multiple recombination events or exposure of cryptic modifier loci. See S1 Table. Complementation of a69 Recombinant Strains.

The top panel shows the cytogenetic map of the 51 region of chromosome 2R [74], used with permission. Diagramed below are the deleted regions of deficiency strains tested, the corresponding molecular map and identified protein coding genes in the region. Arrows or arrowheads indicate the orientation of gene transcription and arrow or arrowhead length corresponds to gene length at the scale indicated (K, kilobase). Data obtained from Flybase version FB2018_01 [64].
To identify the gene specifically in the a69 mutation, we took two approaches. First, a subset of genes were examined for alterations in expression in the a69 mutant, and second, a large series of complementation studies were performed with alleles of known mutants in the region. cDNAs from 5 genes in the region were obtained and in-situ hybridization of third instar larval eye imaginal discs was performed on cn1 bw1 (wild-type) and a69 mutants. In each case the expression pattern of the gene was not substantially disrupted in a69 mutants, suggesting that the phenotype is not due to the disruption of patterned mRNA expression of these genes in the 3rd instar eye-antennal disc. (Fig 5). hibris (hbs) was expressed strongly in the morphogenetic furrow and maintained weakly posteriorly, consistent with a previous report [27]. It was also expressed in the ocellar region and in the developing antenna. parcas (pcs, FBgn0033988) was expressed strongly in the morphogenetic furrow and in the antenna. CG10265 (FBgn0033990) did not appear to be expressed in either the eye or antennal regions. CG7639 (FBgn0033989) appeared to be weakly expressed in the region anterior to the morphogenetic furrow. caskin (ckn, FBgn0033987) was expressed anterior to the furrow and in the antenna.

The figures shows in situ hybridization of biotinylated reverse strand probes prepared from hibris, parcas, CG10265, CG7639, and caskin cDNA clones (rows) against wild type (cn1 bw1) (left column) or a69 mutant (right column) eye-antennal imaginal discs.
We characterized Rh5 and Rh6 expression in animals heterozygous for a69 and alleles of Additional sex combs (Asx, FBgn0261823), atypical protein kinase C (aPKC, FBgn0261854), bocce (boc, FBgn0011203), charlatan (chn, FBgn0015371), Enhancer of GMR-sina 2-1 (ES2-1, FBgn0024358), Hexokinase C (Hex-C, FBgn0001187), knot (kn, FBgn0001319), Regulatory particle non-ATPase 6 (Rpn6, FBgn0028689), safranin (sf, FBgn0003367), Protein 1 of cleavage and polyadenylation factor 1 (Pcf11, FBgn0264962), scab (scb, FBgn0003326), and transposon insertions P{A26O9}1 (FBti0001751) and P{lacW}B6-2-25 (FBti0005748). All of these mutations complemented a69 (data not shown).
We obtained the following alleles of hbs: hbs361(FBal0130217), hbs459, (FBal0130216), hbs1130(obtained from M. Baylies) and hbs2593 (FBal0130218). With one exception, all of these alleles fail to complement a69, Table 2. Furthermore, hbs361 homozygotes and heteroallelic combinations of these alleles all show a substantial decrease in the proportion of Rh5 expression in R8 photoreceptor cells. With four exceptions, viable combinations of these alleles over deficiencies in the region show the same complementation pattern as the a69 mutant, S3 Table.
Complementation crosses of a69, hbs alleles and cn bw control.
Exon sequencing of the hbs gene failed to identify unique polymorphisms in the a69 mutant that were absent in phenotypically wild type control strains (data not shown). Nonetheless, given that the gene spans over 30 Kb including 24 Kb in the first intron, it seems likely that a mutation within a regulatory region of the gene may be responsible for the hypomorphic a69 phenotype and the complex complementation pattern found with the hbs459 allele. Thus, we believe the complementation data is fully consistent with a69 being a hbs allele, hbsa69.
hibris is expressed in the developing third instar eye imaginal disc
Consistent with previous studies [28], we find that hbs is expressed in the developing third instar eye imaginal disc in preclusters of photoreceptor cells emerging from the morphogenetic furrow and ultimately in all photoreceptor cells, Fig 6. hbs is expressed coordinately with early senseless (sens, FBgn0002573) expression in R8 just posterior to the morphogenetic furrow and this is followed by prospero (pros, FBgn0004595) expression in R7 cells 6-8 rows posterior and cone cells.

The figure shows a confocal microscopy flattened Z-stack series of hibris (hbs) immunolabeling (A) and triple labeling of the same wild type (cn1 bw1) specimen with antibodies against hbs, senseless (sens) and prospero (pros) (B). The morphogenetic furrow has moved from right (posterior) to left (anterior).
hibris is required in the retina for R7 and R8 cell differentiation
To examine the function of hbs in Rh5 and Rh6 expression in R7 and R8 photoreceptor cell patterning, we examined an additional allele of hbs in mosaic flies. We used the ey-FLP driver to generate homozygous mutant clones in the retina and optic lobes of animals that were heterozygous for hbs1130. We used a cell autonomous lethal to generate large homozygous mutant clones and eliminate homozygous wildtype tissue, as described [29]. Fig 7 shows a small heterozygous clone with a single Rh5 expressing R8 cell in an otherwise homozygous mutant retina where Rh3 expressing R7 cells are mispaired with Rh6 expressing R8 cells, demonstrating a phenotype identical to a69.

Large FLP-FRT retinal clones were generated in the eye and optic lobes with ey-FLP. Panel A shows a bright field microscopy image of a single heterozygous clone (red tissue marked with w+) within an otherwise homozygous hbs1130 mutant retina (white tissue). B and C show one R8 photoreceptor cell expressing Rh5 in this heterozygous clone, whereas elsewhere in the retina, only Rh6 is expressed in R8 photoreceptor cells. Frequent mispairing of Rh3 expressing R7 cells and Rh6 expressing R8 cells is also shown. Rh3 (blue), Rh5 (red) and Rh6 (green) expression were detected by confocal microscopy with directly labeled monoclonal antibodies as described in Materials and Methods.
To further refine the spatial requirement for hbs in R7 and R8 photoreceptor cell differentiation and opsin gene expression we compared mutant clones of hbs66 (FBal0239852) [30] generated using ey-FLP and ey3.5-FLP [31]. ey3.5-FLP is a modified form of ey-FLP that efficiently induces clone formation in the third instar larval eye imaginal disc, but not in the lamina or medulla. Fig 6A shows loss of hbs in the retina and optic lobe leads to a dramatic decrease in Rh5 expression and mispairing of Rh3 and Rh6 in adjacent R7 and R8 cells of individual ommatidia, consistent with the results obtained with hbs1130, Fig 5. This is in contrast to Rh3, Rh5 and Rh6 expression in a similarly FRT recombined clone of a wild type chromosome. By comparison, retina specific clones generated with ey3.5-FLP [31] also show a loss of Rh5 expression and mispairing of Rh3 and Rh6. These results indicate that hbs is required in the retina for normal R7 and R8 photoreceptor cell differentiation and opsin gene expression.
Overexpression of hibris is sufficient to disrupt R7 and R8 cell differentiation
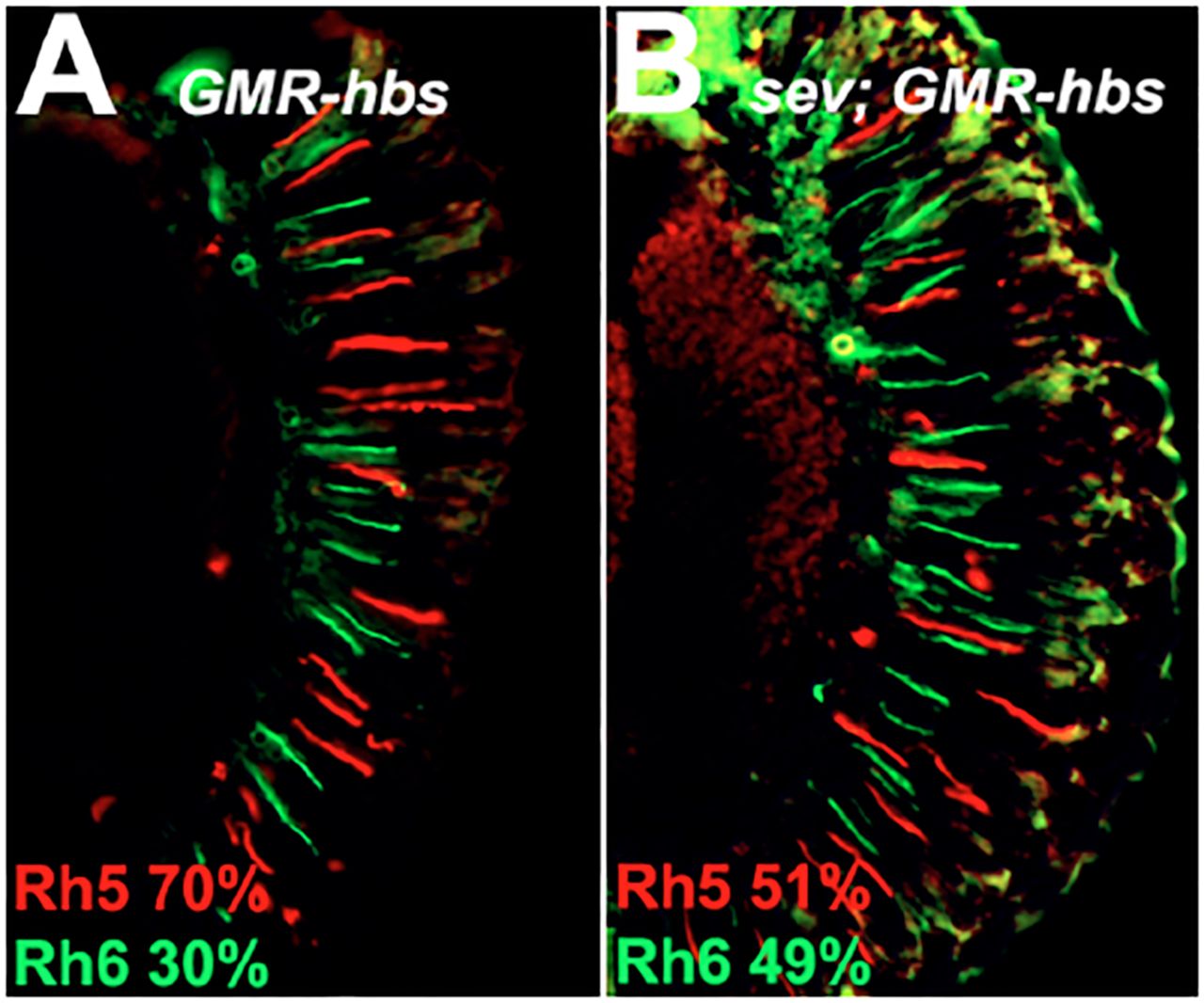
To determine whether ectopic expression of hbs is sufficient to induce the expression of Rh5 in R8 photoreceptor cells, we over-expressed hbs using the GAL4-UAS system [32] and the P{GAL4-ninaE.GMR} driver (FBtp0001315). Fig 7A shows that overexpression of hbs is sufficient to induce Rh5 expression in many, but not all R8 photoreceptor cells. This occurs without perturbation of Rh3/Rh4 R7 cell subtype ratio and is accompanied by mismatched Rh4/Rh5 expressing R7/Rh8 photoreceptor cells pairs (not shown). To test whether the formation of these mismatched ommatidia could result from an inappropriate signal from Rh4 expressing R7 cells or a defect in the default pathway and expression of Rh6 in R8 cells, we overexpressed hbs in a sev mutant background that lacks R7 photoreceptor cells. Fig 7B shows that removal of R7 cells leads to a dramatic reduction but not elimination of Rh5 expression. These results suggest that the ability of overexpressed hbs to induce Rh5 expression in R8 cells is primarily or partially R7 photoreceptor cell independent.
hibris is required non-cell autonomously for R7 and R8 cell differentiation
To determine whether hbs is required cell-autonomously in the R7 and/or R8 photoreceptor cells to enable normal paired expression of Rh3 and Rh5, we generated smaller hbs66 mutant clones in a heterozygous background as previously described [22]. Cells that are either wild type or heterozygous express a myristoylated, membrane associated GFP (myr.GFP, P{GMR-myr.GFP}, FBtp0017435), whereas cells that are homozygous mutant for hbs do not express myr.GFP [33]. We dissociated ommatidia from animals constructed in this manner and counted the expression of Rh5 versus Rh6 in ommatidia that expressed Rh3 in the R7 cell and in which the genotype of the R7 and R8 cells could be scored. Fig 8A shows a cluster of ommatidia from the experiment labeled with antibodies against Rh3, Rh4, Rh5, Rh6, with myr.GFP labeling shown in Fig 8B. Mispairing of Rh3-Rh6 expression occurs in ∼ 20% of cn1 bw1 ommatidia that express Rh3 in R7 cells (Fig 8C), consistent with previous results [11, 22, 23]. In the mosaic analysis, there is a statistically significant increase in the percentage of mispaired Rh3-Rh6 expressing ommatidia ranging from 56 – 100%, regardless of the genotype of the R7 and R8 photoreceptor cells. Particularly noteworthy is the highly abnormal and pronounced effect on ommatidia in which both R7 and R8 photoreceptor cells are heterozygous or homozygous wild-type (R7+ R8+). These results unambiguously indicate that hbs is required non-cell autonomously for the establishment of paired opsin gene expression in the R7 and R8 photoreceptor cells. Although the severity of the mispairing is increased significantly in ommatidia in which R7 and R8 cells are both mutant (R7-R8-).

Large FLP-FRT retinal clones were generated in the eye and optic lobes with ey-FLP, panels A and B, or in the retina alone with ey3.5-FLP. Homozygous hbs66 mutant clones are shown in panels A and C. Homozygous wild-type control clones (+) are shown in panels B and D. Heterozygous tissue is marked with w+ and outlined in panels B, C and D. Rh3 (blue), Rh5 (red) and Rh6 (green) expression were detected by confocal microscopy with directly labeled monoclonal antibodies as described in Materials and Methods.
Notch is required for R7 and R8 cell differentiation
N signaling plays an essential and reiterative role in the development of the compound eye [34], and is known to regulate hbs during myoblast fusion [35, 36], control cell adhesion in the eye through interactions with hbs [37], and hbs may also play a role in N cleavage [28]. To determine whether N is required for the establishment of paired opsin gene expression in the R7 and R8 photoreceptor cells, we reduced N activity at sequential stages of pupal development. We used shifts to a restrictive temperature of a temperature sensitive allele Nl1N-ts1 (FBal0012887) in an otherwise cn1 bw1 background and compared with similarly treated cn1 bw1 controls. Fig 9A shows that at baseline without heat shock Nl1N-ts1; cn1 bw1 flies have a significantly increased proportion of Rh3:Rh4 expressing R7 cells compared to cn1 bw1 controls. The Rh3:Rh4 ratio is significantly increased with heat shock at 24-36 hours after puparium formation (APF) and significantly decreased with heat shock at 36-48 hr APF compared to N mutants raised at the non-restrictive temperature. Because the variation of the Rh3:Rh4 ratio in R7 cells would be expected to alter the Rh5:Rh6 ratio in R8 photoreceptors, we specifically examined the percent of Rh3-Rh6 mispairing as an index of impaired induction of Rh5 expression. We found that Rh3-Rh6 mispairing was significantly increased in both Nl1N-ts1; cn1 bw1 animals and cn1 bw1 control animals heat shocked from 0-12 hr APF, Fig 9B. Statistically significant increases in Rh3-Rh6 mispairing were noted at 24-36 hr APF, 36-48 hr APF and 48-60 hr APF in N mutant animals demonstrating a highly significant disruption in the paired expression of opsin genes in R7 and R8 photoreceptor cells. Because of heat shock induced lethality at 12-24 hr APF in cn1 bw1 control animals, results at this time point are difficult to interpret. However, these results conclusively demonstrate a requirement for N activity in regulating 1) Rh3 versus Rh4 expression in the pR7 and yR7 photoreceptors and 2) coupling of Rh3/Rh5 expression in adjacent pR7 and pR8 photoreceptor cells of the same ommatidium.

Over expression of UAS-hbs with the GMR-GAL4 driver leads to an increase in Rh5 (red) expression, panel A. Removal of R7 photoreceptor cells (sevenless14 mutation) partially suppresses the effect, panel B. Rh6 expression is shown in green.
Materials and Methods
Stocks and Genetics
Stocks were maintained in humidified incubators on cornmeal / molasses / agar media or standard cornmeal food with malt, and transferred on a rotating basis every three weeks as described [38-40]. D. melanogaster strains were obtained from individual laboratories or the Bloomington Drosophila Stock Center (BDSC). Genotypes were constructed using conventional genetic techniques, dominant markers and appropriate balancer chromosomes [39, 41].
Genotypes of Animals Shown in Figures
Figure 2 A, B, C: w1118
Figure 2 D, E, F: w1118; P{etau-lacZ}a69
Figure 5, Left column: WT = cn1 bw1, Right column: w1118; P{etau-lacZ}a69
Figure 6: cn1 bw1
Figure 7: yd2 w1118 P{ry+t7.2=ey-FLP.N}2 / w1118; P{w+mW.hs=FRT(whs)}G13 L* / P{w+mW.hs=FRT(whs)}G13 hbs1130
Figure 8A: w1118/ yd2 w1118 P{ry+t7.2=ey-FLP.N}2 P{GMR-lacZ.C(38.1)}TPN1; P{ry+t7.2=neoFRT}42D hbs66/ P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A l(2)cl-R111
Figure 8B: w1118/ yd2 w1118 P{ry+t7.2=ey-FLP.N}2 P{GMR-lacZ.C(38.1)}TPN1; P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A / P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A l(2)cl-R111
Figure 8C: w1118/P{w+mC=ey3.5-FLP.B}1, y1 w*; P{ry+t7.2=neoFRT}42D hbs66/ P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A l(2)cl-R111
Figure 8D: w1118/ P{w+mC=ey3.5-FLP.B}1, y1 w*; P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A / P{ry+t7.2=neoFRT}42D P{w+t* ry+t*=white-un1}47A l(2)cl-R111
Figure 9A: w1118; P{GAL4-ninaE.GMR}12 / P{UAS-hbs.A}
Figure 9B: w1118 sev14; P{GAL4-ninaE.GMR}12 / P{UAS-hbs.A}
Figure 10: w1118/ yd2 w1118 P{ry+t7.2=ey-FLP.N}2 P{GMR-lacZ.C(38.1)}TPN1; P{ry+t7.2=neoFRT}42D hbs66/ P{ry+t7.2=neoFRT}42D P{w+mC=GMR-myr.GFP}2R

Clusters of dissociated ommatidia from the eyes of hbs66 flies carrying small homozygous mutant clones were examined for expression of Rh3 (yellow), Rh4 (red), Rh5 (blue), Rh6 (magenta) panel A, and myr.GFP (green, panel B). Heterozygous or homozygous wild-type tissue is labeled with a myristoylated, membrane associated GFP (myr.GFP) (green, panel B, labeled R7+ or R8+ in panel C). Homozygous mutant tissue is unlabeled for myr.GFP (labeled R7- or R8- in panel C). Asterisks in panels A and B indicate two ommatidia showing myr.GFP expression in R7, R8 and other (in the case of the left ommatidia in panel B) photoreceptor cells. Panel C shows the percentage of mispairing of Rh3-Rh6 expressing R7-R8 cells in individual ommatidia (Y-axis) for unrecombined cn bw control flies, and each genotype of R7/R8 photoreceptor cells (X-axis). Asterisks (*) in panel C indicate that Rh3-Rh6 expression mispairing is significantly different statistically from the cn bw control (n=96 ommatidia) for R7+ R8+ (p=5.2×10−9, n=170 ommatidia), R7- R8+ (p=1.6×10−6, n=11), R7+ R8- (p=5.0×10−5, n=6), R7- R8- (p<10−15, n=178). Hashtag (#) in Panel C indicates mispairing is significantly different statistically from R7+ R8+ for R7- R8- (p<10−15, n=178). Error bars indicate the 95% confidence intervals for the measured percentages.
Figure 11: cn1 bw1 (cn bw) or y1 Nl1N-ts1 g2 f1; cn1 bw1, (N; cn bw)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nl1N-ts1; cn1 bw1 (shaded columns) flies were compared to cn1 bw1 control (white columns) flies. Panel A shows data for the percent of R7 cells expressing Rh3 versus Rh4. Panel B shows data for the percent of Rh3-Rh6 mispairing in adjacent R7 and R8 photoreceptor cells within an individual ommatidium. Both panels A and B are bar graphs showing no temperature shift (none, raised continuously at 20°C) or a 12 hour temperature shift (temperature raised to 29°C at the indicated time in hours after puparium formation (APF) followed by return to 20°C). Hashtag (#) indicates significantly different from cn bw control receiving the same treatment p<0.05. Asterisk (*) indicates significantly different from the same genotype not heat shocked (none) p<0.05. + not recovered (heat shock induced lethality was observed for cn bw at this temperature shift time point). Error bars indicate the 95% confidence intervals for the measured percentages. Number of ommatidia counted for cn bw and N; cn bw respectively Panel A (no heat shock 259, 446; 0-12 hr heat shock 505, 411; 12-24 hr heat shock 0, 226; 24-36 hr heat shock 285, 306; 36-48 hr heat shock 509, 577; 48-60 hr heat shock 610, 586). Panel B (no heat shock 96, 232; 0-12 hr heat shock 197, 234; 12-24 hr heat shock 0, 104; 24-36 hr heat shock 103, 187; 36-48 hr heat shock 183, 231; 48-60 hr heat shock 232, 316).
Immunohistochemistry
10μm cryosections were prepared and treated as previously described [11]. Dissociated ommatidia were prepared from six animals. Eyes were cut from heads using 28 gauge needles in Phosphate Buffered Saline (PBS). The retina, cornea +/- lamina tissue was shredded with needles, triturated 10 X with a 200 μL pipette tip and transferred to a microscope slide to dry at RT. Subsequent treatment was the same as cryosections. Antibodies were used at the following dilutions: directly conjugated mouse monoclonal anti-Rh5 (Texas Red, 1:100, RRID: AB_2736994) and directly conjugated mouse monoclonal anti-Rh6 (FITC, 1:100 RRID:AB_2736995) [42], mouse monoclonal anti-Rh4 (clone 11E6, 1:10, RRID:AB_2315271) [11, 42], mouse monoclonal anti-prospero (1:10, RRID:AB_528440, [43]), guinea pig polyclonal anti-senseless (1:1000, [44]), rabbit polyclonal anti-hibris (1:400, AS-14, RRID:AB_2568633, [45]). Secondary reagents were obtained from Life Technologies Corporation (Carlsbad, CA) or Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA). An additional reagent was prepared from purified (Cell Culture Company, LLC, Minneapolis, MN) mouse monoclonal anti-Rh3 (RRID:AB_2315270). anti-Rh3 was directly conjugated using Alexa Fluor™ 647 Protein Labeling Kit (Invitrogen, catalogue number A20173) and used at 1:100 dilution. Immunofluorescence images were acquired with an Axioskop plus/AxioCamHRc (Carl Zeiss, Inc., Thornwood, NY) or by confocal microscopy using a Zeiss Pascal LSM (Carl Zeiss, Inc.) or Leica TCS SP5 (Leica Microsystems Inc., Buffalo Grove, IL).
Statistical Analysis
Comparisons of the proportions (percentages) of opsin expression in different genetic backgrounds were performed with a z-score and are shown in Table 1 and Table 2 [46]. The z-score was calculated using the equation:
 p1 and p2 = proportions of marker expression in each of the two different genotypes under comparison. n1 and n2 = number of ommatidia counted for each genotype. pavg = average proportion for both genotypes combined. qavg = 1-pavg. The significance of the difference between the two proportions was determined from the normal distribution as a one- or two-tailed test. The 95% confidence interval of a proportion was calculated using the Wilson procedure without continuity correction [47, 48] using VasarStats [49].
p1 and p2 = proportions of marker expression in each of the two different genotypes under comparison. n1 and n2 = number of ommatidia counted for each genotype. pavg = average proportion for both genotypes combined. qavg = 1-pavg. The significance of the difference between the two proportions was determined from the normal distribution as a one- or two-tailed test. The 95% confidence interval of a proportion was calculated using the Wilson procedure without continuity correction [47, 48] using VasarStats [49].
RNA in situ hybridization
Eye-antennal imaginal discs from third instar larvae were dissected in PBS, fixed in 50mM EGTA / 4% formaldehyde in PBS, rinsed in methanol, and stored in ethanol at -20°. Discs were treated with ethanol/xylene (1:1), rinsed with ethanol, post-fixed in 5% formaldehyde in PBS plus 0.1% Tween (PBT), washed with PBT, and digested with Proteinase K (5 μg/ml). Tissue was post-fixed again and pre-hybridized in hybridization buffer (50% deionized formamide, 5XSSC, 1 mg/ml glycogen, 100 μg/ml salmon sperm DNA, 0.1% Tween) at 48°C. Discs were hybridized overnight at 55°C with 2 μl digoxigenin-labeled antisense RNA probe in 100 μl hybridization buffer. Probes were prepared from cDNA clones D1 [50], GH09755 (FBcl0125531), GM02985 (FBcl014202), LD18146 (FBcl0156485), LP09461 (Fbcl0187603) of genes hbs, pcs, CG10265, CG7639 and ckn, respectively. The hybridized imaginal discs were washed extensively with hybridization buffer at 55°C followed by PBT washes at room temperature. Discs were incubated with alkaline phosphatase-conjugated anti-digoxigenin antibody (1:2000, Roche Applied Science, Indianapolis, IN) overnight at 4°C. Discs were washed with PBT and gene expression was visualized with staining solution (100mM NaCl, 50 mM MgCl2, 100 mM Tris pH 9.5, 0.1% Tween) containing NBT/BCIP (Roche Applied Science). Stained imaginal discs were mounted and photographed using an Axioskop plus/AxioCamHRc (Carl Zeiss Inc.).
Notch Temperature Sensitivity
Stocks of y1 Nl1N-ts1 g2 f1 / C(1)DX, y1 f1; cn1 bw1 or control cn1 bw1 flies were maintained at 20°C. Male offspring were collected at the white prepuparium stage (P0). At 0, 12, 24, 36 and 48-hours after puparium formation (APF), pupae were shifted to 29°C for 12 hours and returned to 20°C until eclosion or formation of pharate adults. Retinas were dissected and ommatidia were dissociated and labeled with opsin antibodies for cell counting. 30 pupae were collected in each experimental or control group.
Discussion
Here we describe the isolation and characterization of a novel allele of the D. melanogaster gene hibris, an evolutionarily conserved NPHS1 (nephrin) related IgSF member [51]. We show that hibris is required for the coordinated expression of opsin genes in adjacent R7 and R8 photoreceptor cells within the compound eye. Orthologues of this gene have been identified in many species, and numerous paralogues within species play diverse roles in organ system development and function [52]. Within the context of R7 and R8 photoreceptor cell differentiation and the regulation of opsin gene expression in the retinal mosaic, the specific functional role of hbs is unclear.
As noted briefly in the Introduction, the current model for the establishment of paired opsin gene expression in the R7 and R8 photoreceptors requires the type I activin receptor baboon (babo, FBgn0011300), bone morphogenetic protein type 1B receptor thickveins (tkv, FBgn0003716), transforming growth factor (TGF) beta type II receptor punt (put, FBgn0003169), many of their ligands, ligand processing convertases, and downstream effector enzymes [21]. In addition, the tumor suppressor kinase warts (wts, FBgn0011739), hippo kinase (hpo, FBgn0261456), salvador (sav, FBgn0053193), and melted (melt, FBgn0023001) a modulator of insulin/PI3K signaling [12], the hpo signaling cascade members Merlin (Mer, FBgn0086384), kibra (kibra, FBgn0262127), and the tumor suppressor lethal (2) giant larvae (l(2)gl, FBgn0002121) [19], and the transcription factors ocelliless (oc, FBgn0004102), dorsal proventriculus (dve, FBgn0020307) [53], PvuII-PstI homology 13 (Pph13, FBgn0023489) [54] and erect wing (ewg, FBgn0005427) [55] are also required. Although not specifically tested in every case, all of these genes are thought to function cell autonomously within the R7 or R8 photoreceptor cells.
hbs is required in the eye for the induction of Rh5 expression based upon our experiments making homozygous mutant clones with ey3.5-FLP (Fig 8). However, in mosaic animals in which the R7 or R8 cells may be mutant or heterozygous in a mixed genotype environment, we find that hbs appears to have both cell autonomous and non-cell-autonomous effects. Specifically, ommatidia that carry R7 and R8 cells that are genotypically homozygous mutant are significantly more likely to show Rh3-Rh6 expression mispairing (Fig 10C, p<10−15). By contrast, in mosaic animals in which both the R7 and R8 cell of an individual ommatidium are heterozygous or homozygous wildtype, there is still a substantial reduction in Rh5 expression in R8 cells and a statistically significant (Fig 10C, p=5.2×10−9) increase in Rh3-Rh6 expression mispairing. This reflects a classic non-cell autonomous effect.
What could that effect be? Traditionally inductive processes are thought to occur between tissues or cells in which there is an inducer and a responder. Inductive signals are also often defined as instructive or permissive [56]. In the presence of an instructive interaction (i.e. from a pR7 cell), the responder (R8) develops in a certain way (as a pR8 cell expressing Rh5). By contrast, in the absence of the instructive interaction (yR7 or R7 cells absent, e.g. seveneless (sev) mutants), the responder (R8) does not develop in a certain way (does not become pR8 expressing Rh5, but rather becomes yR8 and expresses Rh6 instead as a default fate (with some exceptions [11]). If hbs played a formal instructive role in regulating the expression of Rh5 in R8 photoreceptor cells, then we would expect that its expression throughout the retina (GMR-Gal4; UAS-hbs) would lead to expression of Rh5 in all R8 photoreceptor cells even in the absence of R7 cells (Fig 9B). While the number of R8 cells expressing Rh5 is far higher than in sev mutants alone [10, 11, 22, 23], ectopic expression of hbs in this experiment is not sufficient to induce Rh5 expression in all R8 photoreceptor cells. Therefore, hbs does not play a strictly instructive role in this process.
As a potentially permissive regulator of R8 photoreceptor cell differentiation, hbs may play a role in establishing the architecture of the developing eye. Perhaps loss of hbs in mosaic or fully mutant animals disrupts cellular contacts that mediate signaling between R7 and R8. There is ample evidence for disruption of cone and pigment cell differentiation and eye roughening in hbs mutants [57, 58]. Furthermore, hbs and its binding partner roughest (rst) are known to have effects on axon guidance and synapse formation in the optic lobes [59-62]. Perhaps interactions within the lamina or medulla are responsible for some aspect of inductive signaling and expression of Rh5 in pR8. Finally, perhaps the loss of Rh5 expression in the hbs mutant eye reflects an inability to respond to the inductive signal, a loss of competence [63]. We previously suggested that rhomboid (rho, FBgn000463) and the Epidermal growth factor receptor (Egfr, FBgn0003731) may play a role in establishing competence of the R8 cell [22]. In these studies, we showed that rho is required for the induction of Rh5 expression in R8 photoreceptor cells, but like hbs, when rho is lost in mosaic retinas but the R7 and R8 cells of an individual ommatidia are wild type or heterozygous, there remains a dramatic effect on induction of Rh5 expression. Furthermore, loss of Egfr was also found to reduce the induction of Rh5 expression and also affect the proportion of pR7 and yR7 cells. These findings suggest that hbs likely plays a permissive, non-cell autonomous role in R7 and R8 differentiation.
Because hbs is both regulated by N signaling [50] and also thought to participate in N processing following its activation [28], our finding that N is also required for induction of Rh5 expression is not unexpected. This result is completely consistent with the previous findings that sequential loss of N signaling disrupted eye development and differentiation at every time point [34]. Similarly, alterations in N signaling also affect the proportion of pR7 and yR7 cells, suggesting that it as well as Egfr may regulate what is thought to be a cell-autonomous stochastic developmental process. These findings raise several notes of caution regarding the potential complexity of the system and the need to rigorously test the underlying hypotheses upon which the current model for R7 and R8 photoreceptor cell differentiation is based.
Subsequent analysis of the role of hbs in R7 and R8 photoreceptor cell differentiation will require further identification of its specific interaction partners in this system, either in the retina or optic lobes, as well as the temporal requirement for its involvement in R7 and R8 cell differentiation. Ample resources are available including available mutant strains [64], RNAi transgenics [65], and temporal and spatial mis-expression tools [66-70]. Despite these technical resources, defining the precise role of hbs in R7 and R8 differentiation will likely yield a complex system, reflecting coregulation of the IRM proteins [71], involvement of large complexes associated with scaffolding proteins [72], functional or genetic redundancy, compensation [73] and feedback.
Supporting Information
S1 Table. Complementation of a69 Recombinant Strains. Recombinants described in Fig 3 were crossed to a69 and the number of ommatidia counted expressing Rh5 or Rh6, Total counted, and % Rh5 are indicated in the table. Controls for comparison were homozygous a69 mutants or a69 / w1118 heterozygotes. Each recombinant strain was compared to both controls (right two columns) and was either not significantly different (NSD) or significantly different from (SDF) the indicated control at the p value stated. Statistical comparisons of strains were carried out as described in Materials and Methods. Controls are indicated at the bottom of the table. Recombinant strains having % Rh5 values intermediate between wild type and mutant phenotypes, but statistically significantly different from both, are shaded.
S2 Table. Complementation of a69 by Deficiency Strains. A panel of thirty three deficiency stains were crossed to a69 to test for complementation. The number of ommatidia counted expressing Rh5 or Rh6, Total counted, and % Rh5 are indicated in the table. The control for comparison was homozygous a69 mutants. Compared to a69 (right column) each deficiency over a69 was either not significantly different (NSD) or significantly different from (SDF) a69 at the p value stated. Statistical comparisons of strains were carried out as described in Materials and Methods. Values for the a69 mutant are indicated at the bottom of the table. Deficiency strains failing to complement a69, which are not statistically significantly different from a69, are shaded.
S3 Table. Complementation of hibris alleles by Deficiency Strains. A panel of seven deficiencies were crossed to a69, hbs361, hbs459, hbs1130, hbs2593 and cn1 bw1 to test for complementation of the a69 mutant phenotype. The number of ommatidia counted expressing Rh5 or Rh6, Total counted, and % Rh5 are indicated in the table. The control for comparison was homozygous a69 mutants. The deficiencies failed to complement the tested genotype (shaded rows) or complemented the tested genotype (white rows). Complementation was defined as significantly greater Rh5% than (SGT) a69 homozygous mutant at the p value shown using a one-tailed test. Statistical comparisons of strains were carried out as described in Materials and Methods. Values for the a69 mutant are indicated at the bottom of the table. Crosses having results that differed from expected are noted (Exceptions).
S4 Table. Strain Information. Includes recombination stocks, deficiencies and alleles that complement a69. Stock genetics, Flybase ID and RRID are listed where available.
Acknowledgements
We thank Mary Baylies, Ruben Artero, Gerry Rubin, Amy Tang, James Mohler, and Jeff Sekelsky for D. melanogaster stocks, Mary Baylies for the hbs D1 cDNA clone, Karl Fischbach for the rabbit anti-hbs antibody (AS-14), and Hugo Bellen for the guinea pig anti-sens antibody. Stocks obtained from the Bloomington Drosophila Stock Center (NIH P40OD018537) were also used in this study. We thank Natalia Toledo Melendez for technical assistance, John Aldrich and Tom Jacobsen for comments on the manuscript and thoughtful discussion.
References




