

Abstract
Although muscle development has been widely studied in Drosophila melanogaster there are still many gaps in our knowledge, and it is not known to which extent this knowledge can be transferred to other arthropods. To help closing these gaps we participated in a large-scale RNAi screen that used the red flour beetle, Tribolium castaneum, as a screening platform. The effects of systemic RNAi were screened upon double-stranded RNA injections into appropriate muscle-GFP tester strains. Injections into larvae were followed by the analysis of the adult thoracic muscle patterns, and injections into pupae by the analysis of the late embryonic/larval muscle patterns. Herein we focus on the results of our pupal injection screen for late embryonic/larval muscle phenotypes, which covered 8.500 of a total of ∼16.500 genes of the Tribolium genome. Apart from many genes known from Drosophila as regulators of muscle development, a collection of genes previously unconnected to muscle development yielded larval body wall and leg muscle phenotypes. We present the main candidates that remained after being processed through a series of verification and selection steps. Further, we discuss why distinct though overlapping sets of genes are revealed by the Drosophila and Tribolium screening approaches.
Introduction
Muscle development in arthropods has been studied primarily in the dipteran Drosophila melanogaster, whereas much less is known about the regulatory mechanisms guiding muscle development in other arthropod orders. Thus, it is unknown whether more distantly-related arthropods, such as beetles, utilize largely the same processes and mechanisms to make muscles, or whether they differ in important aspects. Clearly, one shared process of holometabolous arthropods such as dipterans and coleopterans is that the musculature of an animal has to be developed twice; first the larval musculature during embryogenesis and second the adult musculature with completely different features during metamorphosis in the pupae.
In Drosophila, many of the genetic control mechanisms guiding the development of the multinucleated larval muscles of the body wall, also known as somatic muscles, have been uncovered during the past few decades. The earliest events in the hierarchy of zygotically active regulatory genes involve the activation of twist and snail in a ventral strip of cells in the blastoderm embryo, which are needed for the specification of the mesoderm and its invagination during gastrulation (Leptin 1991). Tribolium castaneum (Tc) twist, which is one of the few known regulators of mesoderm and muscle development in the red four beetle, fulfills similar early functions as Drosophila twist (Händel et al. 2005; Stappert et al. 2016). In Drosophila and likely in Tribolium, the subsequent spreading of the internalized mesoderm in tight contact with the overlying ectoderm is facilitated by FGF signals from the ectoderm (Wilson and Leptin 2000; Sharma et al. 2015). Upon formation of a mesodermal monolayer spanning the entire dorsoventral extent of the germ band, patterning events that are largely derived from the ectoderm subdivide the mesoderm along the anterior-posterior and dorsoventral axis within each parasegmental unit, which leads to the formation of the anlagen giving rise the somatic, cardiac, and visceral muscles. With regard to the somatic (and cardiac) mesoderm, a major inductive signal is provided by Wingless (Wg) (Baylies et al. 1995; Lee and Frasch 2000; Bodmer and Frasch 2010; Azpiazu et al. 1996; Riechmann et al. 1997; Lee and Frasch 2000). Subsequently, the somatic mesoderm is further subdivided by intersecting signals from the ectoderm, including Wg and Decapentaplegic (Dpp), into domains that are competent to respond to localized and temporally-regulated receptor tyrosine kinase (RTK) signals (Frasch 1999; Baylies and Michelson 2001).
These signals are mediated by the Drosophila epidermal growth factor receptor (EGFR) or, alternatively, the fibroblast growth factor receptor Heartless (Htl) and define groups of equivalent cells in terms of muscle formation (Carmena et al. 1998). The antagonistic actions of these RTK signaling activities and of Delta/Notch signaling activities within these equivalence groups ultimately results in the formation of two types of myoblasts within each group: A single muscle progenitor in which the RTK signaling cascade remains active and the Notch signaling cascade is inactive, and several adjacent cells with the opposite activity status of these signaling cascades. The muscle progenitor with high RTK and low Notch signaling activity divides asymmetrically and typically gives rise to two muscle founder cells that are programmed to form a single somatic muscle of a defined identity each. The specific identity of each muscle founder is defined by the expression and functions of defined combinations of so-called muscle identity genes, which are induced downstream of the respective RTKs and cooperating signals, and generally encode members of various families of transcription factors (de Joussineau et al. 2012; Dobi et al. 2015). Conversely, within the neighboring cells lacking RTK activities, high Notch signaling activities induce the expression of the transcription factor encoding lameduck (lmd) gene, which defines these cells as fusion-competent myoblasts that, a priori, are not committed to specific muscle fates (Duan et al. 2001; Ruiz-Gomez et al. 2002).
During the next important event, myoblast fusion, fusion-competent myoblasts fuse sequentially to each muscle founder cell and nascent myotube to generate a specific body wall muscle (Kim et al. 2015; Deng et al. 2017). The recognition and adhesion of the two types of myoblast occurs through the engagement of the immunoglobulin (Ig) domain proteins Sticks-and-stones (Sns) and Hibris (Hbs) on fusion-competent myoblasts with the related Ig domain proteins Kin of irre (Kirre) (aka, Dumbfounded, Duf) and Roughest (Rst, aka, IrreC) on the muscle founder cells. Downstream signaling cascades in both cell types lead to the differential assembly of polymerized actin structures at the prospective fusion site. Most prominently, within the fusion-competent myoblasts the extension of actin-propelled protrusions are thought to cause membrane breakdown and fusion pores upon invading the founder cells (Kim et al. 2015; Deng et al. 2017).
Towards the end and after myoblast fusion, the syncytial muscle precursors form extensions that migrate to the specific epidermal muscle attachment sites and make contacts with them. Several regulators involved in myotube guidance and the establishment of initial contacts have been identified (Schweitzer et al. 2010; Maartens and Brown 2015; Schulman et al. 2015). In many cases this involves the release of Slit proteins from tendon cells, their binding to Robo receptors on the myotubes for proper guidance, and subsequent arrest of migration upon the interaction of Robo with the LRT protein on the membranes of the tendon cells. In other cases, interactions between the trans-membrane protein Kon-tiki (Kon, aka Perdido,), its cytoplasmic partner, Grip (a PDZ domain-containing protein), and the cell-surface protein, Echinoid (Ed), are necessary for targeting muscles to their proper attachment sites and making initial connections. During muscle differentiation, myotendinous junctions and muscle-muscle connections at the attachment sites are stabilized by integrin-mediated adhesions to specific extracellular matrix structures between these cells, as well as by the intracellular linkage of integrin-associated proteins to the cytoskeleton in both muscle and tendon cells (Schnorrer and Dickson 2004; Schweitzer et al. 2010; Maartens and Brown 2015).
Muscle differentiation culminates in the assembly of the sarcomeric apparatus, proper positioning of the myonuclei, and the establishment of neuromuscular junctions (Nose 2012; Volk 2013; Schulman et al. 2015; Lemke and Schnorrer 2017). Two key regulators known to act in muscle differentiation in vertebrates, MyoD and Mef2, are present in Drosophila as single orthologs and regulate muscle differentiation (Michelson et al. 1990; Paterson et al. 1991; Bour et al. 1995; Lilly et al. 1995; Arredondo et al. 2001). nautilus (nau; Drosophila MyoD) is required for the formation and differentiation of a specific subset of larval muscles (Balagopalan et al. 2001). Drosophila Mef2, in addition to functioning in terminal muscle differentiation, has also an essential earlier role in myoblast fusion, likely via transcriptionally activating certain myoblast fusion genes (Sandmann et al. 2006; Brunetti et al. 2015).
Prior to adult muscle development, the vast majority of the larval body wall muscles are histolyzed in early pupae and the adult muscles are built (in most cases from scratch) from stem cell-like cells that have been set aside during embryogenesis and are called adult muscle precursors (AMPs) (Gunage et al. 2017). Many, but not all of the known regulators of larval muscle development are being reutilized during adult muscle development (Gunage et al. 2017). Thoracic AMPs associated with wing and leg discs are being patterned by signals from the epidermal cell layer and thus assume differential muscle fates, such as direct versus indirect flight muscles in case of the wing disc (Sudarsan et al. 2001). The indirect flight muscles differ in their ultrastructure from all other fly muscles in that they exhibit fibrillar, stretch-activated myofibers instead of tubular myofiber organizations. The involvement of the transcription factor Spalt major (Salm) as a master regulator in fibrillar muscle development in the indirect flight muscles is currently one of the few documented examples of conserved regulatory processes during muscle development between Drosophila and Tribolium (Schönbauer et al. 2011).
While this brief synopsis shows that past research has yielded a considerable amount of information on the regulation of muscle development in Drosophila, there still remain many gaps in our knowledge. To fill these gaps, large-scale loss-of-function screens have been performed by RNA interference and classical mutagenesis (Schnorrer et al. 2010; Johnson et al. 2013; Hollfelder et al. 2014; Camuglia et al. 2018). Because each of these methods has its own limitations, such as the delayed action of inducible RNAi in Drosophila embryos and the lack of phenotypes with functionally-redundant genes, we chose to undertake an alternative approach. To identify new components required for normal development of the body wall (somatic) musculature in arthropods, and also to begin to understand the similarities and differences between dipterans and coleopterans in this process, we participated in a large-scale systemic RNAi screen in the red flour beetle Tribolium castaneum, termed “iBeetle” (Schmitt-Engel et al. 2015). Our general strategy was to identify genes with interesting knock-down phenotypes in the somatic musculature in Tribolium and subsequently study the functions of their orthologs in Drosophila in more detail. Herein we describe an overview of this screen and provide a first description of the obtained muscle phenotypes. We include examples of genes from the screen that are orthologous to known regulators of muscle development in Drosophila. Of note, we identified and verified genes in the Tribolium screen that have not been implicated in muscle development in previous research.
Materials and Methods
Tribolium strains
All beetles were kept under standard conditions (Brown et al. 2009) on white wheat flour containing 5% dry yeast at 25 °C and shifted to 32 °C for the experiments. The following Tribolium castaneum stocks were used in this study: San Bernardino (SB), black (Sokoloff et al. 1960), piggyBac pig-19 (pBA19) (Lorenzen et al. 2003), D17Xred (Schmitt-Engel et al. 2015).
Cloning of Tc-Mef2 and Tc-duf/Kirre
To synthesize first strand cDNA the Omniscript RT kit (Qiagen) was used. 5 µg total RNA derived from early embryonic stages were reverse transcribed utilizing oligo(dT) primers and following the manufacturer’s instructions. 1 µl of the cDNA synthesis reaction was subsequently used to amplify 1kb fragments of Tc-Mef2 (TC010850) and Tc-duf (TC002914) by PCR using gene specific primers (Tc-Mef2-F GTTTGATCGGTCCGTGCTAT; Tc-mef2-R GACCGCTCCAGGATATTGAA; Tc-duf-F ACGCGACCAGGAAATATCAC; Tc-duf-R GGAAGCTTGGTTCGGTGTAA). The ∼1kb Tc-duf fragment fully includes the iB_03469 sequences at its 3’ portion. The amplified PCR fragments were gel purified and cloned into the pCR©II-TOPO© vector using the TOPO©TA Cloning© Dual Promoter kit (ThermoFisher Scientific) following the manufacturer’s instructions.
Tc-mef2 and Tc-Duf RNA probe synthesis
To synthesize antisense DIG-labeled Riboprobes, 1 µg of linearized pCR©II-Tc-mef2 or pCR©II-Tc-duf/kirre was in vitro transcribed utilizing the DIG RNA labelling kit (Roche) following the manufacturer’s instructions. The DIG-labelled RNA was purified using the RNA cleanup protocol of the RNeasy Kit (Qiagen). Tribolium fixation and in situ hybridization were performed as described previously (Tautz and Pfeifle 1989; Patel et al. 1994).
Double-stranded RNA preparation and injections for RNAi
To synthesize dsRNA of Tc-mef2 and Tc-Duf, 1 ng of pCR©II-Tc-mef2 or pCR©II-Tc-Duf/Kirre were used in a PCR reaction using T7 and T7-SP6 primers. The amplified fragments were purified using the QIAquick Gel Purification Kit (Qiagen) and dsRNA was produced as described in (Bucher et al. 2002). To induce parental RNAi 1 µg/µl of Tc-mef2 or Tc-Duf dsRNA were injected into adult females as described in van der Zee et al. (2006).
The procedure of the iBeetle larval and pupal RNAi injection screen and the procedure for the analysis of late embryonic/early larval muscles are described in detail in Schmitt-Engel et al. (2015). The dsRNAs used in the iBeetle Screen were obtained from Eupheria Biotec GmbH (Dresden). To generate the dsRNAs for the rescreen cDNA was generated using the Transcriptor First Strand cDNA Synthesis Kit (Roche). The cDNA was then used to amplify fragments by PCR with the same primers as used by Eupheria Biotec GmbH (Dresden) for the original iBeetle Screen. The sequences of the dsRNA fragments in the primary screen (iB dsRNAs; annotated in the format iB_nnnnn) and of the dsRNA fragments non-overlapping with the iB fragments (annotated as iB_nnnnn_2) are accessible in http://ibeetle-base.uni-goettingen.de/gb2/gbrowse/tribolium/ (select track “iB dsRNA”). These dsRNAs were synthesized from PCR products using the MEGAscript™T7 Transcription Kit (Ambion).
Research materials and data availability
Materials produced in this study are available upon request. The authors affirm that all data necessary for confirming the conclusions of this article are represented fully within the article and its tables and figures with the exception of sequence information (e.g., for amplification primers) that is available at http://ibeetle-base.uni-goettingen.de/gb2/gbrowse/tribolium/.
Results
Comparison of somatic muscle patterns between Tribolium and Drosophila
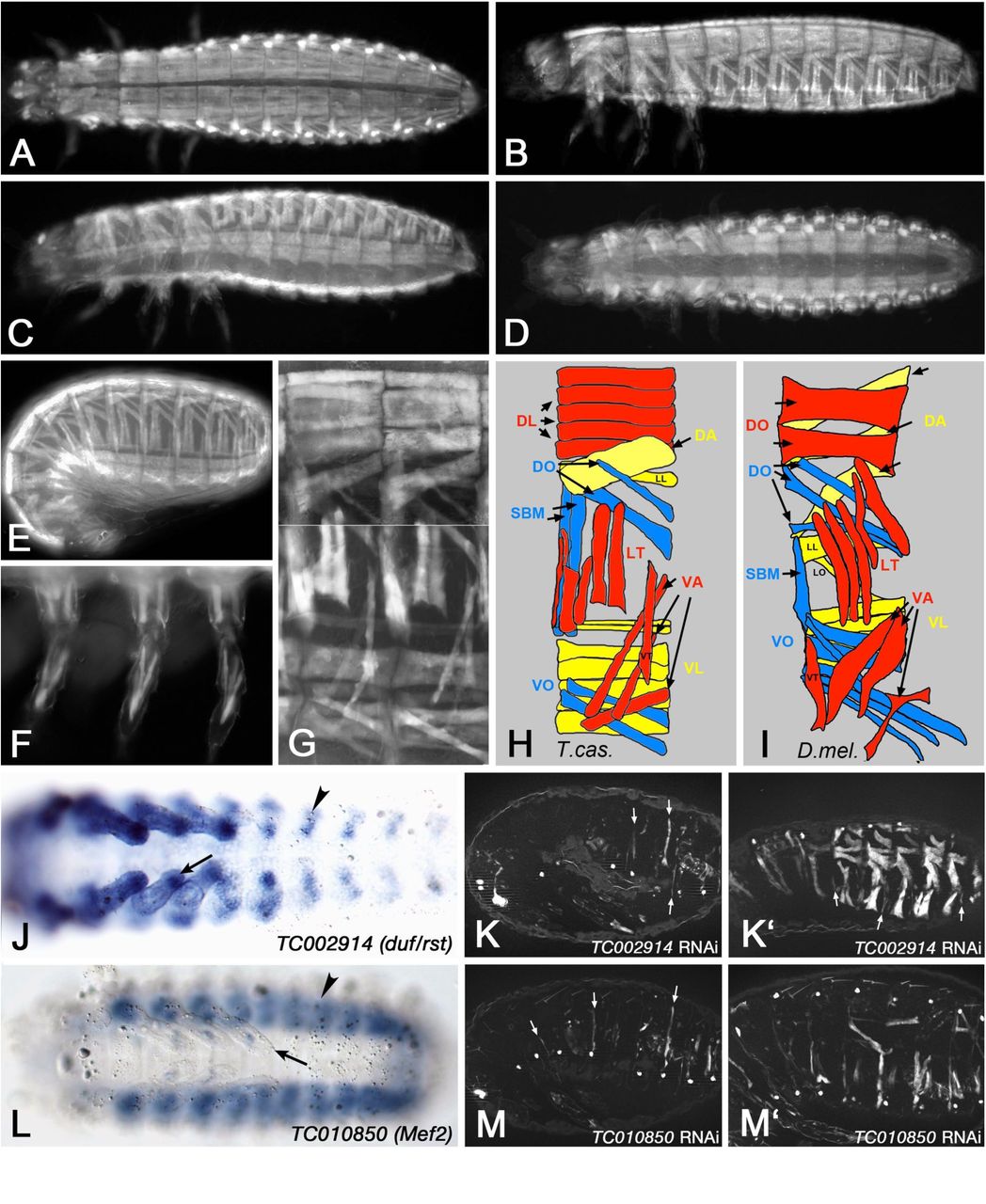
The RNAi screen took advantage of the systemic nature of parental RNA interference in Tribolium castaneum (Bucher et al. 2002). For screening we employed the pig-19 line, which carries a piggyBac insertion in the 3’ UTR of TC003326 (Actin-87E-like) that expresses EGFP in all larval somatic and visceral muscles (Lorenzen et al. 2003). Before screening we used this expression pattern to characterize the wild type pattern of the larval body wall musculature in more detail. As shown in Fig. 1A - E, each abdominal segment displays a stereotypical arrangement of ca. 28 syncytial muscle fibers underneath the body wall, excluding ventral areas where the CNS is located and the dorsal midline where the unlabeled dorsal vessel is positioned. In the thoracic segments, which unlike Drosophila embryos carry appendices, a modified muscle pattern is observed (Fig. 1A - D) and a stereotypic muscle pattern is also seen in the legs (Fig. 1F). The muscle arrangement in each abdominal segment into dorsal, lateral, and ventral groups, as well as their orientations as longitudinal, oblique, transverse, and acute muscles within these groups, are strongly reminiscent of the well-characterized pattern of abdominal body wall muscles in Drosophila (Fig. 1G - I) (Bate 1993). However, the details differ and due to the current lack of conserved markers for individual muscles or sets of muscles it is presently unclear whether any of these muscles are homologous between the two arthropod species.

(A) to (G) show Tribolium castaneum pig-19 enhancer trap 1st instar larvae and embryos imaged live for EGFP expression in the somatic musculature. (A) Newly hatched 1st instar larva, view of dorsal muscle pattern. (B) Newly hatched 1st instar larva, view of dorsal-lateral muscle pattern. (C) Newly hatched 1st instar larva, view of ventral-lateral muscle pattern. (D) Newly hatched 1st instar larva, view of ventral muscle pattern. (E) Late stage embryo prior to hatching, view of lateral muscle pattern. (F) View of 1st instar larval leg muscle pattern. (G) Newly hatched 1st instar larva, high magnification view of muscle pattern in two abdominal segments (composite of a dorsal-lateral and a ventral-lateral view from two different animals). (H) Schematic representation of late embryonic muscle pattern in an abdominal segment from Tribolium castaneum. Note that the external-to-internal orders of the muscles and the exact numbers of DL and VL muscles are tentative. (I) Schematic representation of late embryonic muscle pattern in an abdominal segment from Drosophila (abbreviations: DA: dorsal acute; DL: dorsal longitudinal; DO: dorsal oblique; LT: longitudinal transverse; SBM: segment border muscle; VA: ventral acute; VL: ventral longitudinal; VO: ventral oblique muscles; for nomenclature see (Bate 1993)). (J) In situ hybridization of Tribolium embryo (early germ band retraction stage) for mRNA of TC002914 (ortholog of Drosophila duf/rst). Arrow head: somatic mesoderm in abdominal segment. Arrow: Somatic mesoderm in leg. (K) Late stage pig-19 embryo from adult female injected with dsRNA for TC002914, imaged live for EGFP. Only few and very thin muscle fibers are present (arrows). (K’) Example of milder phenotype in TC002914 knock-down embryo as in (K), showing residual large muscles, very thin muscles (arrows), and gaps where muscles are missing. (L) In situ hybridization of Tribolium embryo (retracted germ band stage) for mRNA of TC010850 (ortholog of Drosophila Mef2). Arrow head: somatic mesoderm in abdominal segment. Arrow: Somatic mesoderm in leg. (M) Late stage pig-19 embryo from adult female injected with dsRNA for TC010850, imaged live for EGFP. Few muscles are present that are very thin (arrows). (M’) Example of milder phenotype in TC010850 knock-down embryo as in (M).
Additional evidence for similarities in muscle development between the two arthropod species came from the expression and function of several Tribolium castaneum (Tc) orthologs of known key regulators of muscle development in Drosophila. The expression of the early mesodermal regulator Twist in the embryonic mesoderm of Tribolium has been documented extensively (Händel et al. 2005; Stappert et al. 2016), and as expected, RNAi against Tc-twist in pig-19 led to a complete absence of muscles (data not shown, see http://ibeetle-base.uni-goettingen.de). TC002914, the single ortholog of Drosophila kirre (aka duf) and roughest (rst), which in Drosophila are essential for myoblast fusion in a functionally redundant manner, is expressed in somatic mesodermal cells of the body wall and limbs in Tribolium embryos (Fig. 1J). Whereas Drosophila kirre is expressed only in muscle founder cells its paralog rst is expressed in both founder and fusion-competent myoblasts (Ruiz-Gomez et al. 2000; Strunkelnberg et al. 2001). The seemingly broader expression of TC002914 as compared to Drosophila kirre may suggest that the expression of TC002914 is more akin to that of rst in Drosophila. This interpretation is supported by the expression of Tc-sticks-and-stones (Tc-sns) (TC032336), which is similar albeit slightly narrower in the somatic mesoderm as compared to Tc-kirre/rst (data not shown). Potentially, Tc-sns mRNA is restricted to fusion-competent myoblasts like Drosophila Sns, which regulates myoblast fusion upon interaction with Kirre. Importantly, RNAi knock-down upon injections of Tc-kirre/rst dsRNA into in pig-19 adult females led to strong reductions in both numbers and sizes of GFP-stained muscles (Fig. 1K, K’), which corresponds to analogous phenotypes in kirre rst double mutants in Drosophila (Ruiz-Gomez et al. 2000; Strunkelnberg et al. 2001). In the most severe examples, only few small muscle fibers were present (Fig. 1K), whereas in milder cases presumably resembling partial knock-downs a fraction of the muscles were present but many others were very thin or missing (Fig. 1K’). Incidentally, this example also illustrates that RNAi screens in Tribolium have the potential to identify myogenic regulators that may have been missed in Drosophila screens due to the presence of functionally-redundant paralogs, if these have only a single ortholog in the beetle. Another example of similarities in myogenic regulation between the two arthropod species is provided by Mef2, which in Drosophila is expressed in all muscle progenitors (and muscles) and encodes a crucial muscle differentiation factor (Lilly et al. 1994; Nguyen et al. 1994). In Tribolium embryos, the Mef2 ortholog TC010850 is also expressed broadly in the somatic mesoderm of the body wall and the limb (Fig. 1L). Like with Drosophila Mef2 mutant embryos, knock-down of TC010850 upon dsRNA injections into adult females caused almost complete loss of muscles as detected by GFP in pig-19 embryos (Fig 1M, M’) (Bour et al. 1995; Lilly et al. 1995). Additional examples of phenotypes of genes orthologous to known regulators of Drosophila myoblast fusion are shown in the accompanying paper (Schultheis et al., 2018b).
Regimen of screening and verification of candidate genes
The main screen (‘pupal injection screen’), which we focus here upon, involved injections of dsRNAs (named iB RNAs) for a total of ∼ 8,500 genes into the body walls of female pig-19 pupae. After crossing the eclosed females (if viable and fertile) with black males the GFP-marked muscle patterns of their offspring were analyzed live under a fluorescence microscope in late embryonic stages and newly hatched first instar larvae, as shown for controls in Fig. 1. The screen was performed in two consecutive rounds and included screenings for various additional phenotypes by the screening consortium, as was described in an overview of the results of the first screening round of ∼5,300 genes (Schmitt-Engel et al. 2015). A parallel screen (‘larval injection screen’) involved injections of a total of ∼5,000 iB dsRNAs into L6 stage larvae of the D17Xred strain and the analysis of the GFP-marked late pupal thoracic muscle patterns (as well as additional phenotypes), as described in Schmitt-Engel et al. (2015).
In the first-pass screen with pupal dsRNA injections, 229 of the ∼8,500 tested genes were annotated with specific embryonic muscle phenotypes upon iB dsRNA injections that were not deemed to be secondary to broader disruptions such as segmentation defects, severe embryonic malformations, early developmental arrest, etc. (Fig. 2). In the first-pass larval injection screen, 96 of the ∼5000 tested genes were annotated with specific pupal muscle phenotypes, only six of which were also found in the pupal screen. In the following, we focus particularly on the genes with embryonic muscle phenotypes detected in the pupal injection screen. As shown in Table 1, 24 of these corresponded to genes with Drosophila orthologs that have been implicated in various aspects of Drosophila muscle development. Although in this first-pass screen most of the muscle phenotypes were not characterized and annotated in detail, the phenotypes for the knock-downs of several Tribolium genes were reminiscent of the muscle phenotypes of mutations in their orthologs in Drosophila. For example, knock-down of the Tribolium ortholog of the Drosophila muscle identity gene org-1 led to the absence of specific muscles, including the segment border muscles like in Drosophila (Fig. 3B, cf. Fig. 3A) (Schaub et al. 2012). Likewise, knock-down of the ortholog of nautilus (nau; Drosophila MyoD) led to a loss of muscles and reduction of the GFP differentiation marker, although the observed phenotype appears more severe as compared to Drosophila nau mutants (Balagopalan et al. 2001) (Fig. 3C). Also knock-down of the kon-tiki (kon) ortholog caused the absence of subsets of muscle fibers, which in Drosophila kon mutants is attributed to defects in myotube migration and attachments in subsets of muscles (Schnorrer et al. 2007) (Fig. 3D). Knock-downs of the Tribolium orthologs of inflated (if) (Fig. 3E) and stripe (sr) (Fig. 3F) led to the appearance of spherical myotubes. This is likely due to disrupted muscle attachments because of weakened integrin-mediated adhesions with tendon cells (in the case of if) or the absence of differentiated tendon cells (in case of sr), as shown previously in Drosophila mutants of their respective orthologs (Brown 1994; Volk and VijayRaghavan 1994). A smaller number of muscle regulatory genes known from Drosophila showed phenotypes in Tribolium late pupal muscles upon knock-downs in the larval injections screen (Table 1 and data not shown). In sum, among the Tribolium orthologs of 76 Drosophila genes connected with various roles in muscle development, 28 showed a knock-down phenotype in late embryonic or (more rarely) in late pupal muscles. For 26 genes no such phenotype was annotated in the first pass screen, 8 were not screenable for muscle phenotypes due to lethality, sterility, or broad embryo disruptions prior to muscle formation, and 14 have not been screened yet by dsRNA injections (Table S1). These data, as well as the data for muscle phenotypes upon knock-downs of genes previously not implicated in muscle development can be accessed in the searchable iBeetle database (http://ibeetle-base.uni-goettingen.de), along with other morphological defects that were screened for in these large-scale RNAi screens (Dönitz et al. 2015; Dönitz et al. 2018).
Compilation of screening data for Tribolium orthologs of Drosophila genes known to regulate various aspects of muscle development with no annotated muscle phenotypes, uninterpretable muscle phenotypes, or no injected dsRNAs.
Compilation of screening data for Tribolium orthologs of Drosophila genes known to regulate various aspects of muscle development with clear Tribolium muscle phenotypes (from primary screen; see also http://ibeetle-base.uni-goettingen.de)
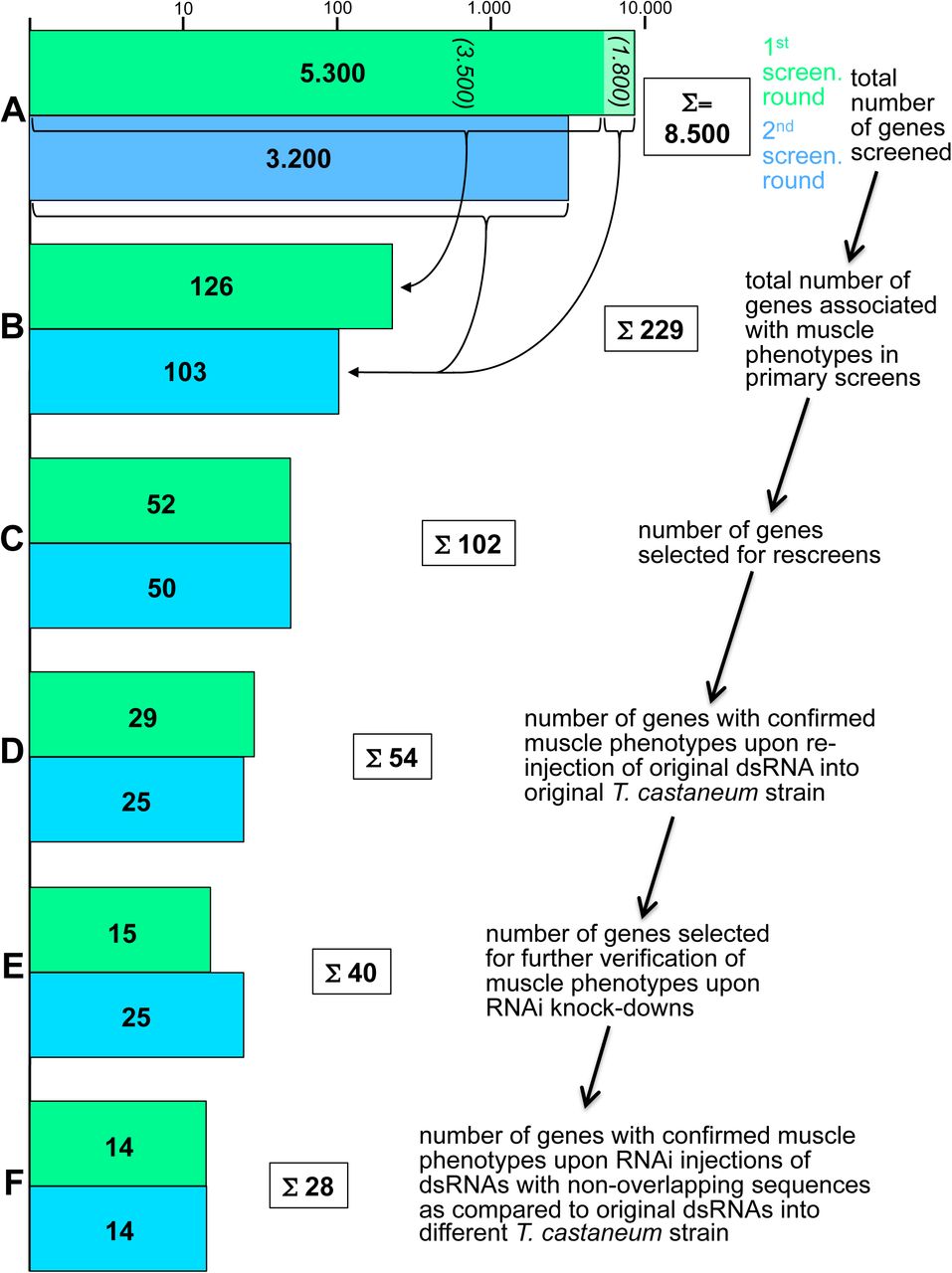
Green bars show numbers of screened and selected genes in the first round and blue bars the corresponding numbers in the second round of screening (blue-green bars in (B) to (F) include “stragglers” from the first-round) (logarithmic scale). (A) A total of 8.500 genes were screened by dsRNA (“iB fragment”) injections in the two rounds. (B) 126 out of 3.500 injected dsRNAs from the first round and 103 out of another 1.800 injected dsRNAs from the first round and from 3.200 of the second round were annotated with muscle phenotypes in the primary screens. (C) 102 from the 229 genes in (B) were selected for rescreens. (D) 54 of the 102 genes from (C) were confirmed for embryonic muscle phenotypes upon re-injection of the original iB dsRNA fragments into pig-19 pupae. (E) 40 of the 54 genes from (D) were selected for independent verification of the observed muscle phenotypes. (F) 28 of the 40 genes from (E) showed confirmed muscle knock-down phenotypes upon pupal injections of dsRNAs non-overlapping with the original iB dsRNA fragments (“NOFs”) into a different T. castaneum strain, San Bernardino (SB).

{kind=link}
{kind=link}
{kind=link}
Shown are lateral or dorsal-lateral views of late stage pig-19 embryos prior to hatching, live imaged for EGFP. (A) Control embryo from uninjected female pupa. DO: Dorsal oblique muscles; SBM: segment border muscles. (B) Embryo with RNAi knock-down of TC015327 (Tc-org-1). Arrows indicate areas of missing or strongly reduced SBM muscles. (C) Embryo with RNAi knock-down of TC015855 (Tc-nau/MyoD). (D) Embryo with RNAi knock-down of TC004764 (Tc-kon). Arrows indicate areas of missing DO muscles. (E) Embryo with RNAi knock-down of TC001667 (Tc-if). (F) Embryo with RNAi knock-down of TC004846 (Tc-sr).
In the next step, we performed a rescreen with 102 of the 229 iB dsRNAs that had annotated muscle phenotypes in the first-pass screen in order to confirm these phenotypes. Because of our aim to identify new Drosophila genes with functions in myogenesis we omitted the genes that have Drosophila orthologs with known roles in myogenesis and those that lack Drosophila orthologs (except for a few with striking and highly penetrant phenotypes). In addition, iB dsRNAs that produced phenotypes with very low penetrance were omitted, as were those that upon closer inspection of the database were likely to yield indirect effects on muscle development. In this rescreen, again performed in pig-19, the muscle phenotypes were confirmed for 54 of the original iB dsRNAs (Figure 2). Particularly in the first round of first pass screening, it turned out that many of the false-positives were due to GFP leakage from muscles upon injury of the late stage embryos or larvae during preparation and mounting. In the second round of first pass screening this effect was taken into account.
In a second rescreen, 40 of the corresponding genes were tested again with the aim to exclude off-target effects and possible strain specific effects. To reduce the work load, 14 genes were omitted, including some that showed only maternal expression in Drosophila or others that encoded enzymes with potentially broader or “house-keeping” functions. These included for example TC010977 and TC002552 that encode an elongase of very long fatty acids and a cytochrome P450, respectively. However we note that genes of this type may still have interesting functions in muscle tissues (see Wang et al. 2016; Xu et al. 2018). For the 40 selected genes, new dsRNAs with sequences that did not overlap with the original iB dsRNA sequences were injected into female pupae from the San Bernardino (SB) strain, and the muscle patterns were analyzed in embryos from a cross of these females with pig-19 males. In addition, the original iB dsRNA fragments were tested by analogous SB pupa injections. These tests served to rule out off-target effects and to confirm an essential role in muscle formation in different genetic backgrounds. As a result, the knock-down phenotypes in the embryonic musculature were confirmed for 28 of the 40 genes with the non-overlapping dsRNAs (Fig. 2). Their phenotypes and additional information on the affected genes are shown in Table 2. These genes show a broad spectrum of distinct muscle phenotypes and the encoded proteins belong to a variety of different protein classes, most of which have not been implicated in muscle development in Drosophila.
Features and muscle phenotypes of selected genes and confirmed in rescreens with original (iB) and non-overlapping (NOF) dsRNA fragments Table 2A Features and muscle phenotypes of selected genes from 1st screening round and confirmed in rescreens with original (iB) and non-overlapping (NOF) dsRNA fragments
Features and muscle phenotypes of genes selected genes from 2nd screening round and confirmed in rescreens with original (iB) and non-overlapping (NOF) dsRNA fragments
Discussion
The iBeetle RNAi screen for muscle defects in Tribolium complements analogous screens in Drosophila
Using Tribolium as high-throughput RNAi screening platform, we uncovered a number of genes that had not been implicated in muscle development in previous work in Drosophila. Conversely, not all orthologs of known Drosophila muscle genes were recovered (27 out of 62; these Drosophila genes were identified via both forward and reverse genetics). This suggests that the different properties of these alternative screening platforms sometimes reveal different facets of a given biological process.
In Drosophila, several forward genetic screens using fluorescent reporter lines for somatic muscles have been performed. In two screens MHC-tauGFP, which marks all somatic muscles similar to the EGFP enhancer trap line used herein in Tribolium, was employed to screen for late embryonic muscle phenotypes upon ethyl methanesulfonate (EMS) mutagenesis (Chen and Olson 2001; Chen et al. 2008). Although the full screens have not been published, several mutants in genes regulating myoblast fusion, myotube targeting and attachment, and muscle maturation have been recovered from these (Chen and Olson 2001; Chen et al. 2003; Schnorrer et al. 2007; Johnson et al. 2013). Other EMS mutagenesis screens employed a cytoplasmic RFP reporter or a nuclear dsRed reporter driven by the founder cell enhancers of the muscle identity gene org-1 and apterous (ap), respectively, in small subsets of muscles. From the org-1-RFP screen, mutants in the genes for the extracellular matrix proteins laminin β and collagen IV α1 have been reported to date, which revealed important roles of these proteins in muscle attachments, e.g., to the cardiac ECM (Hollfelder et al. 2014). From the ap::NLSdsRed screen, genes such as esconsin (ens) were identified that regulate nuclear positioning within myotubes (Metzger et al. 2012). For a number of reasons, including incomplete coverage of the genome and the possible failure to detect more subtle muscle phenotypes, none of these screens reached saturation. RNAi screens for muscle phenotypes upon gene knock-downs have been performed in Drosophila as well, which used lethality and locomotion or flight behavior as initial screening criteria. Instead of injections with dsRNAs, these screens employed inducible expression of dsRNAs with the UAS/GAL4 system and muscle-specific drivers. Due to the generally low knock-down efficiency of this method in embryonic stages, screening for phenotypes was largely confined to larval, pupal, and adult stages. In a large-scale RNAi screen with Mef2-GAL4 driving transgenic UAS-IR RNAi insertions, initial screening was for lethality and flightlessness, and follow-up analyses involved the examination of sarcomeric GFP markers (Schnorrer et al. 2010). This screen identified a large number of known and yet uncharacterized muscle-intrinsic players acting in muscle morphogenesis and function, including spalt, which turned out to be an evolutionarily-conserved master regulator of fibrillar flight muscle development during metamorphosis in arthropods (Schönbauer et al. 2011). Among the 23 genes with verified Tribolium muscle phenotypes (Table 2) that have Drosophila orthologs covered in the Schnorrer screen, only six were annotated for phenotypes in the primary screen in Drosophila (CG11526: early pupal lethality; ths: weak fliers; sau: semilethality; babo: late pupal lethality; croc: weak fliers; MTA1-like: flightlessness) and none of these were followed up with analyses of muscle phenotypes. Reasons for the viability and normal flight capabilities of the other 17 could include, 1) redundant gene functions in Drosophila, as exemplified by Nostrin in the accompanying paper (Schultheis et al, 2018b), or mild defects not leading to lethality or overt flight defects; 2) divergent functions of the fly orthologs; 3) delayed functional knock-down in Drosophila embryos and absence of post-embryonic function in muscle development; 4) ineffective inverted repeat RNAs in Drosophila or false-positives in Tribolium in spite of the verification steps taken. In a recent small scale screen of 82 genes for larval locomotion defects, four genes with orthologs positive in the iBeetle screen were included: twi and Vrp1, which caused lethality, if which caused increased larval locomotion upon knock-down, and nau which lacked any locomotion phenotype. singed (sn, Drosophila fascin), which decreased locomotion, was shown to affect myoblast fusion to a similar degree as our Nost cip4 double mutants (Camuglia et al. 2018; Schultheis et al, 2018b). Tc-sn (TC006673) was not annotated with a muscle phenotype in the iBeetle screen.
The iBeetle screen for genes with knock-down phenotypes in the somatic muscles had the technical advantage provided by the systemic agency of RNAi in Tribolium (Bucher et al. 2002), which made the injection work more economical due to the ease of injecting larvae and pupae and the recovery of large numbers of offspring from each injected female animal. In addition, this procedure is expected to knock down both the maternal and zygotic contributions of genes, and thus has the potential to uncover genes that may have been missed in the Drosophila screens due to maternal rescue. In contrast to the mesoderm-specific RNAi screens in Drosophila but similar to the EMS screens, the iBeetle RNAi screen involved global knock-downs of gene functions. Hence, in addition to genes with muscle-intrinsic functions, genes with non-autonomous functions in muscle developments could be recovered as exemplified by stripe (Fig. 2), which in Drosophila is essential for muscle attachments through its function in determining epidermal tendon cell fates (Becker et al. 1997; Vorbrüggen and Jäckle 1997). On the other hand, global knock-downs increase the risk of secondary effects on muscle development, e.g., due to early effects of genes on developmental events prior to muscle development such as embryonic patterning, cell proliferation, etc‥ These effects were minimized because the iBeetle screen included careful analyses of the larval cuticles with the aim to recover patterning genes as well (Schmitt-Engel et al. 2015; Ansari et al. 2018). Thus, even if some of the genes on our shortlist (Table 2) have additional roles in other tissues, their most prominent functions are expected to be in the development of the somatic musculature, either via mesoderm-intrinsic or via non-autonomous mechanisms.
As for any other screen, it is clear that we missed many genes affecting Tribolium muscle development. Among the Tribolium orthologs of 62 genes known to affect Drosophila muscle development that were included in the iBeetle screen, 27 were recovered through their knock-down phenotypes in the musculature. This is a surprisingly low portion given that the positive controls of genes with a variety of developmental functions included in the screen indicated a detection rate of about 80-90% (Schmitt-Engel et al. 2015). Further analyses are required to test whether this is due to false negative annotations in the screen or to biological differences in the muscle developmental program between the two arthropod species. One potential reason for missing larval muscle phenotypes in false-negatives could be the relatively long time delay between the pupal injections and the onset of embryonic muscle development, which in some cases may reduce the dsRNA concentrations below a critical threshold. As indicated by the rather severe phenotypes obtained with adult injections of Tc-duf dsRNAs as compared to those with pupal injections for several other myoblast fusion genes, adult injections may sometimes provide stronger effects. We did not systematically explore this possibility because pupal injections were necessary for other participants in the consortium, who screened for ovary and oogenesis defects (Schmitt-Engel et al. 2015). As in other screens with pan-muscle markers, it is also likely that some genes with more subtle knock-down phenotypes, e.g., affecting individual or small subsets of muscles, were missed and indeed, the majority of recovered phenotypes affected muscles globally (for an exception, see TC009963, Table 2).
Novel genes identified in the iBeetle screen
Of note, many Tribolium genes were recovered for which their orthologs were not known to affect muscle development in Drosophila. A few genes lack any orthologs in Drosophila although some of them do have orthologs in vertebrates. A notable example for the latter is Tc-Rbm24 (TC001720), for which knock-downs exhibited severe muscle phenotypes (Table 2; Schmitt-Engel et al. 2015). Tc-Rbm24 mRNA is specifically expressed in the developing and mature embryonic somatic musculature and, more weakly, in the dorsal vessel (as well as in the CNS; DS and MF, data not shown). During our screen, mouse RNA-binding motif protein 24 (Rbm24) (and presumably its paralog Rbm38 (Miyamoto et al. 2009)) was reported to play a major role in embryonic skeletal muscle development by regulating alternative splicing of a large number of muscle-specific primary transcripts (Yang et al. 2014). It is conceivable that in Drosophila the role of Tc-Rbm24 in muscle development is exerted by other members of the RRM superfamily of RNA binding proteins. Starting to explore this possibility, we found that some Drosophila RRM superfamily members are expressed in the embryonic somatic mesoderm (boule, CG33714, Hrb87F; in the case of boule exclusively so; DS and MF, data not shown). In addition to Tc-Rbm24, three other genes encoding putative Tribolium RNA binding proteins were recovered (TC010637, TC006055, TC010693; Table 2), reinforcing the important contribution of RNA metabolism in regulating normal muscle development (see also examples from Drosophila; Volk et al. 2008; Johnson et al. 2013; Oas et al. 2014; Spletter et al. 2015).
Several other identified genes shown in Table 2 encode predicted chromatin regulators, which are likely to influence gene regulatory programs during muscle development (TC005276, TC005276, TC006419, TC009963) and possibly have additional, perhaps less prominent roles in other tissues. Tc-croc (TC002813, Table 2), which encodes a forkhead domain transcription factor, exclusively affects the ventral muscles, suggesting that it may act as a muscle identity gene in Tribolium. Interestingly, Drosophila croc appears to be expressed in subsets of ventral mesodermal cells during early muscle development, but we have been unable to detect any ventral muscle defects in Drosophila croc mutant embryos (Häcker et al. 1995; MW and MF, unpublished data).
The Drosophila and mammalian counterparts of TC032839 protein, Unc-76 and FEZ1, respectively, bind to the Kinesin-1 Heavy Chain (KHC). FEZ1 binding in combination with JNK interacting proteins (JIP1, and perhaps similarly JIP3) was shown to release Kinesin-1 autoinhibition and thus activate the motor protein for microtubule binding and motility (Blasius et al. 2007; Koushika 2008). In accordance with this molecular interaction, Drosophila and C. elegans unc-76 were shown to be required for axonal outgrowth and transport (Bloom and Horvitz 1997; Gindhart et al. 2003). In Drosophila kinesin and kinesin-associated proteins, including JIP1/Aplip1 and JIP3/Synd, were shown to regulate nuclear positioning within muscle syncytia (Metzger et al. 2012; Schulman et al. 2014; Auld et al. 2018). Therefore we presume that Unc-76 likewise is involved in this regulatory pathway. Tc-Unc-76 and Dm-Unc-76 are both expressed in the somatic mesoderm (and more prominently in the CNS, as well as maternally), but unlike with RNAi in Tribolium, CRISPR/Cas9-generated zygotic null mutants did not show any overt muscle morphology phenotype in Drosophila embryos (DS, MW, and MF, unpublished data). Therefore, future analyses should investigate myonuclear positioning in Drosophila mutants that lack both the maternal and the zygotic contributions of Unc-76.
In addition, three signaling components were identified, namely Tc-fgf8, Tc-Babo, and Tc-Pvf3. TC-fgf8 (TC000278) encodes the single Tribolium FGF8 member and is a putative ligand for the single, mesodermally-expressed FGF receptor Tc-fgfr, both of which were recently shown to be required for maintaining the expression of Tc-Twist in the somatic mesoderm of late stage embryos (Sharma et al. 2015). This requirement, along with a possible requirement for the activation of yet undefined differentiation genes, could explain the complete absence of pig-19 muscle GFP in Tc-fgf8 RNAi embryos. It will be interesting to investigate in more detail how far muscles can develop in the absence of FGF8 signals. Tc-babo (TC003240) encodes a Tribolium TGF-beta type 1 receptor. The thin-muscle phenotype upon knock-down is reminiscent of phenotypes obtained upon knock-downs of myoblast fusion genes (Schultheis et al., 2018b) and in this case could perhaps be due to under-proliferation of the fusion-competent myoblasts. Tc-Pvf3 (TC008417) encodes a putative ligand for the Tribolium PDGF/VEGF related receptor. In Drosophila this ligand/receptor interaction is required for normal proliferation, migration, and maintenance of hemocytes (Parsons and Foley 2013; Sopko and Perrimon 2013). Hemocytes, in turn, are required for the deposition of extracellular matrix components in the Drosophila body wall (Matsubayashi et al. 2017). Thus it is conceivable that the phenotype of rounded muscles seen upon Tc-Pvf3 knock-down is due to muscle detachments as a result of deficient extracellular matrix at their attachment sites (Maartens and Brown 2015). Following up on these avenues, and likewise on the exact involvement of the other genes identified in the screen, could yield unexpected insights into new aspects in the regulation of arthropod muscle development (for additional preliminary and unpublished data see Schultheis, 2016 (in German)).
Conclusion
The iBeetle RNAi screen has identified numerous genes that are candidates for regulators of arthropod muscle development. In many cases, their orthologs were already known to play roles in muscle development in Drosophila, and in some cases in vertebrates. A significant number of the genes identified in Tribolium had not been recovered in previous Drosophila work before. It will be interesting to examine the roles of identified genes previously not implicated in muscle development in detail in both Tribolium and in Drosophila. In Tribolium, the functional studies by RNAi can now be complemented by CRISPR/Cas9 induced mutations and engineered gene loci (Gilles et al. 2015). In parallel, more detailed analyses of the process of Tribolium muscle development will provide interesting insight into the similarities and differences of muscle development in beetle as compared to fly embryos. In addition, the Drosophila orthologs of newly identified genes with muscle phenotypes in Tribolium can be studied for potential functions in fly muscle development and added to the well-developed framework of regulatory networks in this system. The accompanying paper (Schultheis et al., 2018b) presents an example of this approach by showing that the Drosophila ortholog of the F-Bar domain encoding gene Tc-Nostrin, identified in the iBeetle screen through its muscle phenotype, together with related F-Bar proteins plays a role in Drosophila myoblast fusion and the morphogenesis of adult midgut muscles. Functional redundancy in the fly previously had impeded the identification of this role in Drosophila.
Acknowledgments
We acknowledge support by the German Research Foundation (DFG) for the iBeetle project (FOR1234). We thank Michael Schoppmeier and Martin Klingler for their vital contributions to the organization of the screening at Erlangen.
References




