

Abstract
Neuroimaging studies on language have revealed that the conjunction between speech production and comprehension elicited a common implication of frontal and temporal regions and it is now admitted that speech perception and production rely on both auditory and motor areas. This overlap has been revealed to be also present between reading and listening, but so far no study has simultaneously investigated the overlap of regions involved in the three language tasks (production, perception and reading). The aim of this study is to identify leftward heteromodal brain areas common to these 3 language tasks and to question the existence of right brain areas. Using functional MRI and a new functional regional atlas totally suited for investigating brain lateralization, we performed a conjunction of activated and asymmetrical brain areas in 144 right-handers, to determine the core language areas involved in word list production, perception and reading. Results revealed 3 leftward-clusters, the first one included areas along the central sulcus and premotor supplementary area, the second one included hand motor area, the third one included two regions of the supramarginal gyrus. One the right, the first cluster out of 3 was made of the superior temporal sulcus (STS3).
Leftward frontal and precentral areas together with temporo-parietal areas have been revealed to be commonly activated and asymmetrical, leading to the hypothesis of the involvement of phonological action perception circuits such as the phonological working memory loop, in which articulatory gestures are the central motor units on which word perception, production and reading would develop and act according to the motor theory of speech. On the right hemisphere, the common activation and asymmetry of the STS3, which is a prosodic integrative area, could reflect the processing of fine spectral details over time.
Introduction
Language is one of the most important and specific cognitive abilities of human beings. According to Saussure (Saussure, 1975), language is a universal structure encompassing the abstract, systematic rules and conventions of a unifying system, which is independent of individual users, whereas speech is the personal use of language, thus presenting many different variations such as style, grammar syntax, rhythm, pronunciation etc. The first phase of language acquisition, or speech acquisition, in new-born babies is perceptual as the infant hears others’ vocalizations; this phase is crucial considering the inability to learn spoken language or even babble normally when infants are born deaf (Oller and MacNeilage, 1983) or in the case of wild children (Curtiss, 1977). The second phase is production, through the process of imitation, highlighting the strong dependency between perception and production. Production also plays an important role in language development as evidenced by persisting difficulties in speech production encountered by infants who were tracheotomized at the time at which they should have normally babbled (Locke and Pearson, 1990). Interestingly, infants discriminate and generate all possible sounds their articulatory system allows to produce (Kuhl, 2004). In fact, language-specific abilities start earlier for perception/discrimination than for production, but once language-specific abilities in production are mastered, the discrimination of foreign sounds decline (Canevari et al., 2013). There exist two divergent theories about the relation of speech perception and production to language. The first one, coined the horizontal view, proposes that the elements of speech are sounds that rely on two separate processes (one for speech perception, the other one for speech production) which are not specialized for language until a cognitive process connects them to each other and then to language (Fodor, 1983). The second theory, coined the vertical view (or motor theory of speech perception), posits that speech elements are articulatory gestures serving both speech perception and production processes which are immediately linguistic, thus requiring no cognitive process (Liberman and Whalen, 2000). At the cerebral level, in line with the motor theory of speech perception, the existence of a bilateral dorsal-ventral model of speech processing, with a preferential leftward involvement, has been widely admitted (Binder et al., 1996; Hickok and Poeppel, 2004; Rauschecker and Tian, 2000). This model has posited: (i) a dorsal pathway, i.e., the “where stream,” where an acoustic-phonetic-articulatory transformation linking auditory representations to motor representations is reported to occur in superior temporal/parietal areas and ultimately in frontal areas (Buchsbaum et al., 2001); and (ii) a ventral pathway, i.e., the “what stream”, where speech-derived representations interface with lexical semantic representation, reported to involve the superior, middle, and inferior temporal gyri (Binder et al., 2000; Hickok and Poeppel, 2000). Interestingly, concerning the dorsal pathway, the postulate of the existence of an auditory-motor system (Hickok and Poeppel, 2000), has been supported by studies aiming at examining the role of motor areas in speech perception. Hence, an fMRI study has revealed that listening to syllables and producing the same syllables lead to a common bilateral network encompassing a superior part of the ventral premotor cortex, suggesting the existence of a common phonetic code between speech perception and production (Wilson et al., 2004). Furthermore, another study has not only put forward that the cortical motor system is organized in a somatotopic way along the precentral cortex, the lip area being superior to the tongue area, but has also revealed that these precentral regions are consistently activated by syllable articulation and syllable perception, hence demonstrating a shared speech-sound-specific neural substrate of these sensory and motor processes (Pulvermüller et al., 2006). These findings have been supported by a meta-analysis revealing that in right-handers, activations of the posterior part of the frontal lobe distributed along the precentral gyrus are strongly left lateralized during both production and auditory tasks (Vigneau et al., 2006). Though mastered more belatedly and through an effortful learning, human beings have developed ways of using language through other sensory modalities, such as the visual system in the case of reading. Different studies examining word processing cerebral networks common to the auditory and visual modalities have revealed the supramodal implication of anterior regions (supplementary motor area – SMA -, prefrontal, premotor and inferior frontal gyrus) whereas variations have been observed in the temporal lobe (Booth et al., 2002a, 2002b; Buckner et al., 2000; Chee et al., 1999). One study aiming at investigating the impact of modality and linguistic complexity during reading and listening tasks (Jobard et al., 2007) has revealed that the precentral gyrus was one of the supra-modal regions exhibiting a gradual increase of activation depending on complexity (from words to texts). Taken together, these results would suggest the existence of supra-modal brain areas, located in the precentral gyrus and inferior frontal gyrus involved in the phonological loop, common to three language tasks, i.e., speech production, speech perception and reading, which has not been investigated so far. Furthermore, as the brain functions in networks, it could be hypothesized that in line with this frontal activation, other components of the auditory motor loop, such as the superior marginal gyrus (SMG), or temporo-parietal areas such as the Sylvian-parietal-temporal area (Spt), which belongs to the phonological loop (Buchsbaum et al., 2011), would also share a common recruitment for the three language tasks. Even if language, and more particularly, articulation is left lateralized, it is well established that the right hemisphere is involved in the prosodic dimension of speech (Beaucousin et al., 2007; Belin et al., 2004; Sammler et al., 2015). To assess the hypothesis of a bilateral neural recruitment, it appears more suitable to use lists of over-learnt words, such as months of the year, making it possible to decrease the weight of lexico-semantic and syntactic processing while inducing a prosodic processing due to the specific metrics of lists. Furthermore, brain areas which are not devoted to language but to the performance of the tasks per se, such as monitoring, selecting, keeping the instructions in mind should also share a bilateral common recruitment encompassing areas belonging to executive and visual processing.
Moreover, handedness should be taken into account since language has long been reported as being left lateralized in right-handers, the production task being the most lateralized language task (Binder et al., 2011; Mazoyer et al., 2014; Stroobant et al., 2009; Tzourio-Mazoyer et al., 2015). Indeed, evidence from dichotic listening paradigms (Kimura, 1961) and aphasia (Dax, 1865) point to a left-hemisphere (LH) dominance of language processing. Studies on split-brain patients have also supported the idea of a leftward lateralization concerning phonological processing (Gazzaniga, 2000). Indeed, split-brain patients’ right hemisphere lacks categorical perception of phonemes (Sidtis et al., 1981) and is unable to demonstrate systematic phonology (Gazzaniga, 2000). Studies with the Wada test procedure have also confirmed the leftward-asymmetry of the audio-motor loop, which is the specific support of the phonological processing of speech dimension (Dym et al., 2011). Hence, it appears of great importance to take into account brain asymmetries that is to say compare activation in homotopic areas. Since the brain presents a global torsion, the Yaklovian torque, which makes difficult a point-to-point correspondence between cortical areas that are functionally homotopic (Toga and Thompson, 2003), the use of flipped images appears problematic since it cannot ensure that the flipped regions correspond to the equivalent of the other hemisphere. A new atlas AICHA, based on resting state fMRI data and composed of homotopic functional Regions Of Interest (hROIs), has been devised to circumvent this latter problem and is thus suited for investigating brain hemispheric specialization and lateralization, allowing for determining right and left hemispheric contribution in language and for computing functional asymmetries in regions having equivalent intrinsic connectivity (Joliot et al., 2015). Hence, combining activated and asymmetrical brain areas, makes it possible to assess brain lateralization and to determine the core language areas involved in word list production, perception and reading.
The aim of the present study was twofold. (1) To identify the leftward-hetero-modal brain regions common to three language tasks, i.e., speech production, speech perception and reading with a comprehensive investigation of how these conjointly activated and asymmetrical areas variations are coloured by the task to permit an elaboration on their function/role (2) To apply the same methodology to question the existence of right brain areas devoted to word processing independently of the task since right areas are involved in paralinguistic analysis such as prosodic and emotional processing.
Based on a large population, this study aimed at identifying the regions showing the combined left activation and asymmetry in the 3 tasks on the one hand and the combined right activation and asymmetry on the other end. Due to inter-individual variability of hemispheric asymmetries, 144 right-handed subjects were selected from the BIL&GIN database (Mazoyer et al., 2014), since, even when showing lower lateralization, right-handers have a left hemisphere dominance for language as assessed by the production task both at the group level (Tzourio-Mazoyer et al., 2016), and at the individual level (Zago et al., 2017).
Material and methods
Participants
The present study included a sample of 144 right-handers balanced for sex (72 women) from the BIL&GIN database, which is a multimodal imaging/psychometric/genetic database specifically designed for studying the structural and functional neural correlates of brain lateralization (Mazoyer et al., 2016). Participants were selected as having French as their mother tongue and were free from developmental disorders, neurological and psychiatric history. A local ethic committee (CCPRB Basse-Normandie, France) approved the experimental protocol. Participants gave their informed, written consent, and received an allowance for their participation. All subjects were free of brain abnormalities as assessed by an inspection of their structural T1-MRI scans by a trained radiologist.
The mean (± standard deviation) age of the sample was 27 years ± 6 years (range: 19–53) and the mean level of education (corresponding to the number of schooling years since the first grade of primary school) was 16 ± 2 years (range: 11–20) corresponding to four years at the university level.
Handedness was self-reported by the subjects and their Manual Lateralization Strength (MLS) was assessed using the Edinburgh inventory (Oldfield, 1971) whose values ranged from −100 to +100. Average MLS values were 93.48 (S.D. = 11.49) for the subjects of the present study.
Manual ability was quantified using the Finger Tapping Test (FTT) completed by each participant. Subjects had to hit a button with the left or right index finger as many times as possible during ten seconds while maintaining their wrist on the table. Each measure was repeated three times, and results were averaged for each hand. An asymmetry index (FTTasym) was computed as follows: (RFT - LFT / (RFT + LFT) * 100, where RFT and LFT represented the right and left hand finger tapping scores, respectively. Values ranged from −6.2 to 39.4, average FT-Tasym being 7.5 (S.D. = 5.4).
Functional imaging
Paradigm of the word-list tasks
Language mapping was assessed in the three language tasks in which they had to covertly generate (PRODWORD), listen (LISTWORD) or read (READWORD) lists of words. These tasks were part of a run that alternated these word tasks with sentence tasks.
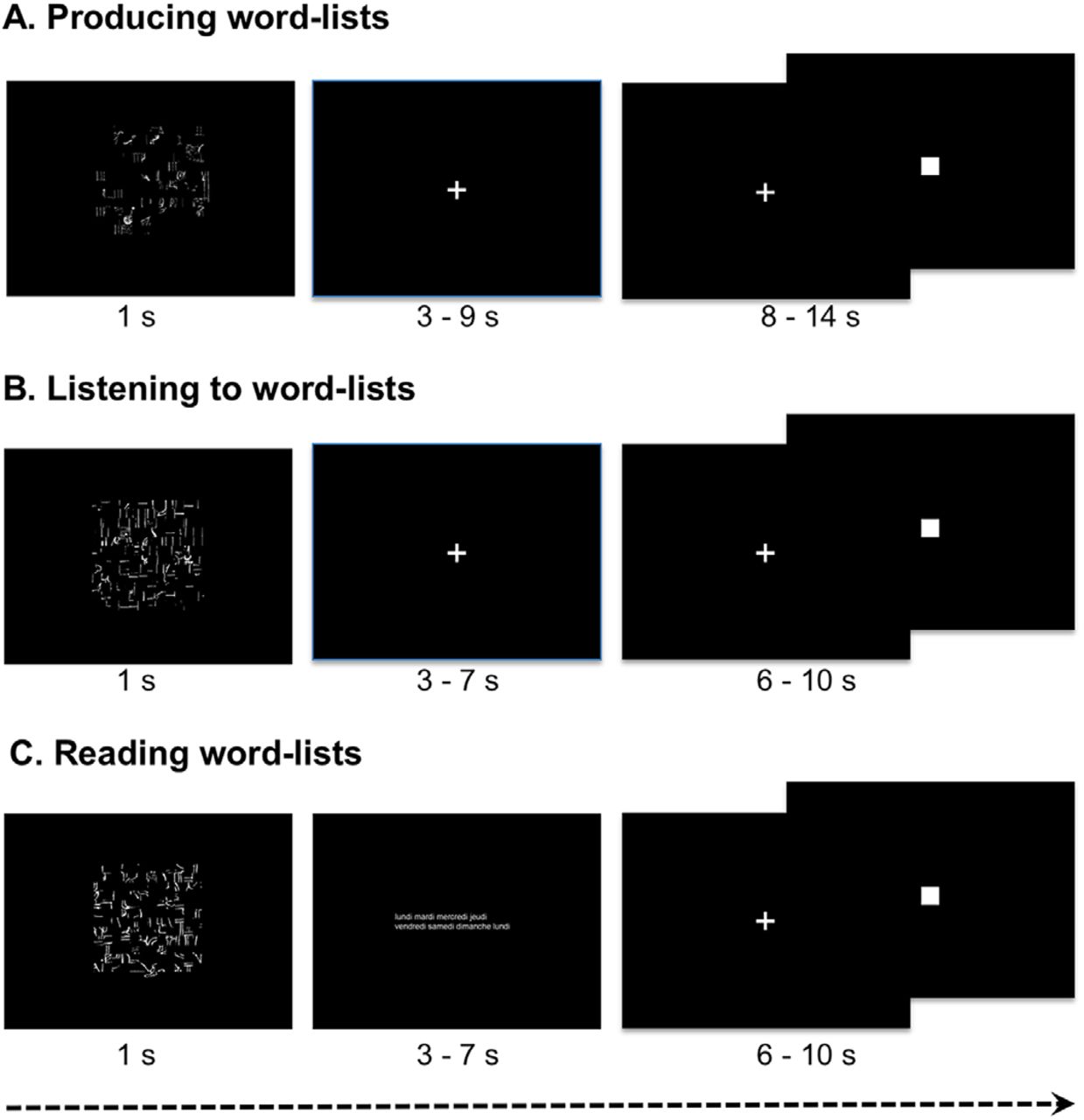
The three language tasks based on word-lists presented the same methodology concerning the experimental paradigm making it possible to not only reveal brain areas common to the performance of the language tasks, but also to the administration of the tasks (Figure 1).

Description of the paradigm for one event of each language task. In the three conditions, an event started by the presentation during one second of a scrambled picture that was followed by a central cross the participants were instructed to fixate while they were producing the list of months of the year (A), or listening to a list of words (B). During the reading run instead of a cross fixation, lists of words (either weeks, hours, seasons, days, months) were presented. They had to click when they had finished to produce, listen to or read and they had next to indicate by clicking when the central cross that reappeared changed into a square.
They were presented as following:
For each task, subjects were shown a scrambled drawing during 1 s, immediately followed by a central fixation crosshair. While fixating the cross, the subjects performed either the listening task or the production task. During the reading task, subjects were asked to read lists of words, which were flashed on the screen. Once the task was completed, a low-level reference task, which consisted in sustaining visual fixation on the cross and pressing the pad when the fixation cross was switched to a square, was presented. The square was then displayed up to the end of the trial. This second part of the trial, which lasted at least half of the total trial duration, aimed at refocusing the subject’s attention to a non-verbal stimulus and at controlling for the manual motor response activation, which was also present in the first part of the trial. A 12-s presentation of a fixation crosshair preceded and followed the first and last trial, respectively. Note that except during the scrambled drawings display, the subjects were asked to keep fixating the cross, and then the square presented at the center of the screen. The drawing size covered an area of 14×14 degrees of visual angle. The fixation cross, the star and the square sizes covered 0.8×0.8 degrees.
For each task, subjects had to fixate the screen for at least 3 min.
Speech production task
For PRODWORD, when a scrambled drawing was displayed, the subject was asked to covertly generate the list of the months of the years from January to December. This condition, though automatic, involves phonetic encoding, phonology, articulation, word retrieval and prosody. The paradigm consisted in a ten 18-s trial of word-List production. The time limit for response, indicated by the transformation of the cross into a star, was 9 s including the 1-s drawing display. Depending on the generation time varying from a subject to another, the duration of the star presentation was at least 4 s and that of the square was at least 5 s, the total event lasting 18 s. The entire experimental run lasted 6 min, 1 min 30 s being devoted to word-list processing.
Speech perception task
For LISTWORD, when a scrambled drawing was displayed, the subject was instructed to listen to the list of the months, days of the week and/or seasons. The paradigm consisted in a thirteen 14-s trial of wordList listening. The time limit for response, indicated by the transformation of the cross into a star, was 7 s. Depending on the reaction time varying from a subject to another, the duration of the star presentation was at least 3 s and that of the square was at least 4 s before a new trial. The entire experimental run lasted 6 min 4 s, 1 min 31 s being devoted to word-list listening.
Reading task
For READWORD, when a scrambled drawing was displayed, the subject was instructed to read the list of months, days of the week and/or seasons. Participants were asked to read each word at a quiet pace to avoid them to quickly extract the global structure of the list without actually reading it. It is thus likely that their reading included subvocal articulation. The paradigm consisted in a thirteen 14-s trial of Word-List reading. The time limit for response, indicated by the transformation of the cross into a star, was 7 s. Depending on the reaction time varying from a subject to another, the duration of the star presentation was at least 3 s and that of the square was at least 4 s before a new trial. The entire experimental run lasted 6 min 4 s, 1 min 31 s being devoted to word-list reading.
Image acquisition and analysis
Anatomical and functional images acquisition
Imaging was performed on a Philips Achieva 3 Tesla MRI scanner. Structural MRI protocols consisted in a localizer scan, a high resolution 3D T1-weighted volume (sequence parameters: TR = 20ms; TE = 4.6ms; flip angle = 10°; inversion time = 800ms; turbo field echo factor = 65; sense factor = 2; matrix size = 256×256×180; 1mm3 isotropic voxel size) and T2*-weighted multi-slice images were also acquired (T2*-weighted fast field echo (T2*-FFE), sequence parameters: TR = 3,5s; TE = 35ms; flip angle = 90°; sense factor = 2; 70 axial slices; 2mm3 isotropic voxel size). Functional images were acquired with a whole-brain T2*-weighted echo planar images acquisition (T2*-EPI, sequence parameters: 240 volumes; TR = 2s; TE = 35ms; flip angle = 80°; 31 axial slices; 3.75mm3 isotropic voxel size) covering the same field of view than the T2*-FFE acquisition.
Functional imaging analysis
For each participant, (1) the T2*-FFE volume was rigidly registered to the T1-MRI; (2) the T1-MRI was segmented into three brain tissue classes (grey matter, white matter, and cerebrospinal fluid; and (3) the T1-MRI scans were normalized to the BIL&GIN template (aligned to the MNI space), including 80 volunteers from the BIL&GIN database gender-matched using the SPM5 “segment” procedure with otherwise default parameters.
For each of the 3 fMRI runs, data were corrected for slice timing differences. To correct for subject motion during the runs, all the T2*-weighted volumes were realigned using a 6-parameter rigid-body registration. The EPI-B0LD scans were then registered rigidly to the structural T2*-FFE image. The combination of all registration matrices allowed for warping the EPI-BOLD functional scans to the standard space with a single trilinear interpolation.
The regional BOLD values were measured from the first level PRODWORD, LISTWORD and READWORD individual contrasts in pairs of hROIs of the AICHA atlas (Joliot et al., 2015). Among the 192 hROIs of AICHA per hemisphere, 8 were not included because more than 20% of their volume was not mapped in all participants either on the right or on the left, leading to a total of 184 hROIs pairs allowing the calculation, for each pair, of left, right, and left minus right asymmetry.
Statistical analyses
Identification of the hROIs co-activated and co-asymmetrical in the 3 language tasks
In this set of 184 hROIS, a conjunction analysis was conducted by selecting the hROIS having both a significant group-level activation and asymmetry in the 3 tasks. Meaning that the significant threshold for the group-level activation or asymmetry was 0.05/184 = 0.00027, Bonferroni corrected for the 184 tests. As the group-level activation or asymmetry was composed of 3 tasks, and in order to conserve the initial group-level threshold, the final significant threshold for activation or asymmetry for one task was set to (0.00027)1/3 = 0.064 (cubic root of 0.00027).
This procedure was conducted separately for the left and the right hemisphere.
These statistical procedures were conducted using the JMP 11 Pro software package, (www.jmp.com, SAS Institute Inc., 2012).
Investigation of the different profiles of activation and asymmetry within the hROIs selected as co-activated and co-asymmetrical in the 3 language tasks
In a second step, we characterized the regions identified as co-activated and co-asymmetrical in the 3 language tasks (either leftward or rightward), according to their profiles of activation and/or asymmetry according to the 3 tasks.
Identification and characterization of cluster of regions having close profile of activation and asymmetry
We applied an agglomerative hierarchical cluster analysis (aHCA) with Ward’s method to group regions of a given hemisphere having comparable profiles of activity and asymmetry. To select a set of features to describe each hROI, we used the averaging activation matrix: a given hROI being defined by its activations and asymmetries in each task (PRODWORD, LISTWORD and READWORD). An hROI was therefore characterized by these 6 values.
The number of sub-clusters was determined through the R library “NbClust” (Charrad et al., 2014). This package provides 30 statistical indices for determining the number of clusters and proposes the best clustering scheme from the different results obtained by varying all combinations of number of clusters for the chosen methods; here aHCA with Ward’s method. We chose the number of clusters that responded to the maximum of indices.
Post-Hoc analysis was conducted on the mean values of activation and asymmetry of each identified cluster to assess potential differences across tasks.
Reliability of the identified clusters through the population and robustness of the identified clusters with respect to the clustering method
To assess the reliability of each cluster within the population studied, we used multiscale bootstrap resampling (Efron et al., 1996). In order to do so, 10,000 multiscale bootstrap resampling datasets, including 50% to 140% of sample data from the 144 participants (Suzuki and Shimodaira, 2004), were processed. Applying the R package “pvclust” (Suzuki and Shimodaira, 2006) function to the multiscale bootstrap resampling outputs, we measured the approximately unbiased (AU) p-value for each cluster. The AU p-value for a cluster is the probability of this cluster to occur among the 144 participants, and indicates the cluster reliability.
We also assessed the robustness of the clustering method by comparing the aHCA using Ward distance method with 3 other clustering methods, i.e. aHCA using average distance method, the k-Means method and the Gaussian mixture model. Hence, the adjusted Rand index was calculated for each comparison.
Results
Tasks performances during the scanning session
PRODWORD: the mean response time taken for the generation of the months of the year was 5261ms, ± 1092ms, (range: 2836–7360).
LISTWORD: the mean response time taken for listening to the months of the year, days and seasons was 486ms, ± 101ms, (range: 282–794), it is noteworthy that it is shorter than in the 2 others tasks since it was calculated after the delivery of the stimulus, i.e., after 4386ms.
READWORD: the mean response time taken for reading the months of the year, days and seasons was 4405ms, ± 603ms, (range: 2703–5681). There is no significant correlation between the reaction time and the FTT index (R2: 0.003, p: 0.51).
Regions co-activated and co-asymmetrical during reading, listening and production of word-lists
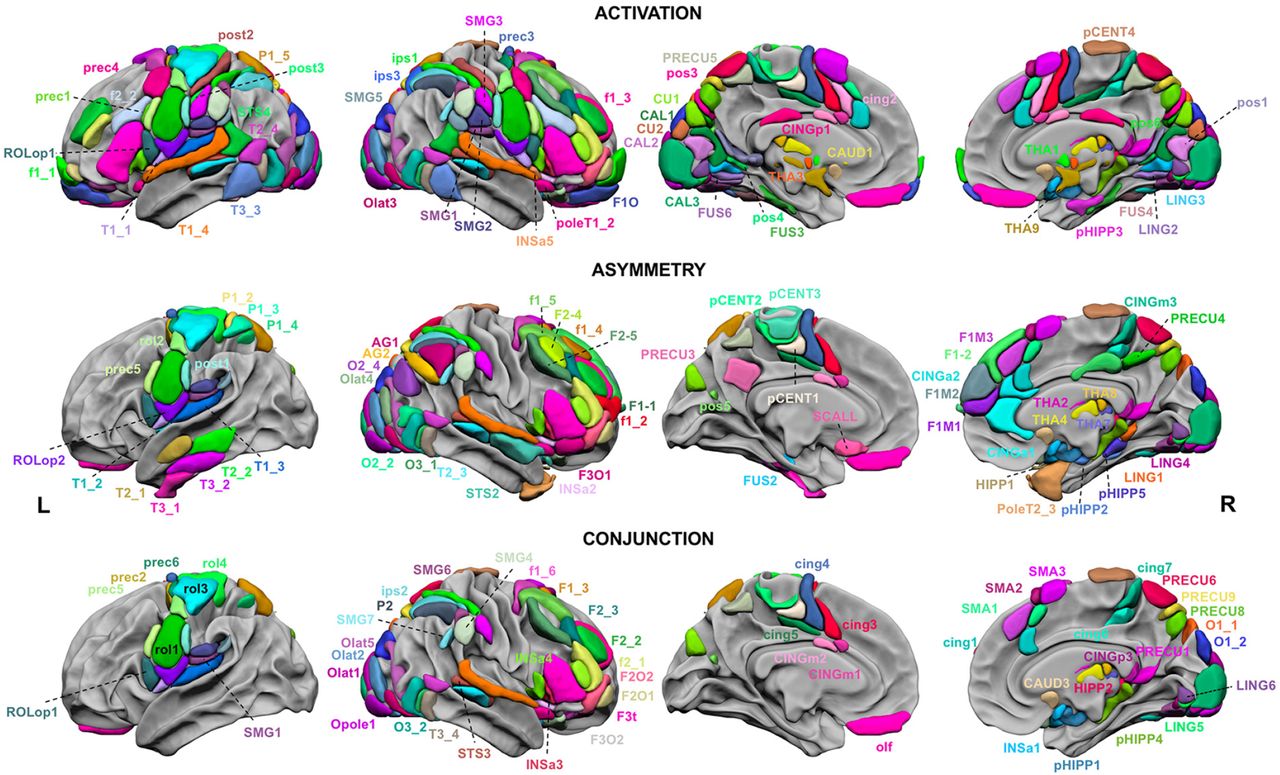
First observation is that there were 15 hROIs conjointly activated in the left hemisphere and leftward asymmetrical while there were only 9 hROIs that were conjointly activated in the right hemisphere and rightward asymmetrical demonstrating the left hemisphere dominance of brain areas dedicated to word-list processing (Figure 2).

Regions of the AICHA atlas significantly activated in the 3 tasks (first row), significantly asymmetrical in the 3 tasks (second row), and significantly co-activated and co-asymmetrical in the 3 tasks (third row). The hROIs are projected on the white matter surface of the BIL&GIN template with Surf Ice (https://www.nitrc.org/projects/surfice/) software. Leftward asymmetrical as well as co-leftward activated and co-asymmetrical hROIS are presented on the left hemisphere and rightward asymmetrical hROIS are presented on the right hemisphere. Selection of hROIs considered as activated or asymmetrical per hemisphere was done according to Bonferroni (p corrected <.00027, 184 hROIs tested), the statistical threshold applied for conjunction of asymmetry and activation for a given task was p corrected =.016 (Bonferroni correction per region and per task, square root of .00027) for each hemisphere and the threshold set for the 3 tasks conjunction was p=.064 (cubic root of .00027, Bonferroni correction).
Left hemisphere
Conjunction of regions showing significant leftward activation and asymmetry in the three tasks
The conjunction of significant leftward activation in the three contrasts (intersection of PRODWORD, LISTWORD and READWORD) revealed 55 left hROIs (Figure 2, first row). The conjunction of significant leftward asymmetry in the 3 contrasts evidenced 27 hROIs (Figure 2, second raw) and 15 left hROIs were commonly activated and leftward asymmetrical (p<.0002, Figure 2 third row, table 1). Most of these hROIs were located in the frontal cortex, including 7 straddling along the Rolandic sulcus and precentral gyrus and 4 located the internal surface of the frontal lobe. Two hROIs were located in the parietal lobe, in the supramarginal gyrus. Finally, one hROI was located in the cuneus and one in the pallidum.
Number of hROIs having significant left activation, leftward asymmetry or conjunction of activation and asymmetry for the 3 word list tasks. b. Number of hROIs having significant right activation, rightward asymmetry or conjunction of activation and asymmetry for the 3 word list tasks. For a given contrast or asymmetry and for a given hemisphere the statistical threshold was set at 0.00027 (Bonferroni correction for 184 hROIs). For the conjunction between activation and asymmetry the statistical threshold was set at 0.016 (Bonferroni correction for the conjunction of two contrasts) and for the conjunction between 3 tasks the statistical threshold was set at 0.064 (Bonferroni correction the conjunction of 3 tasks). The number of regions at a non-corrected threshold of 0.05 is given in brackets.
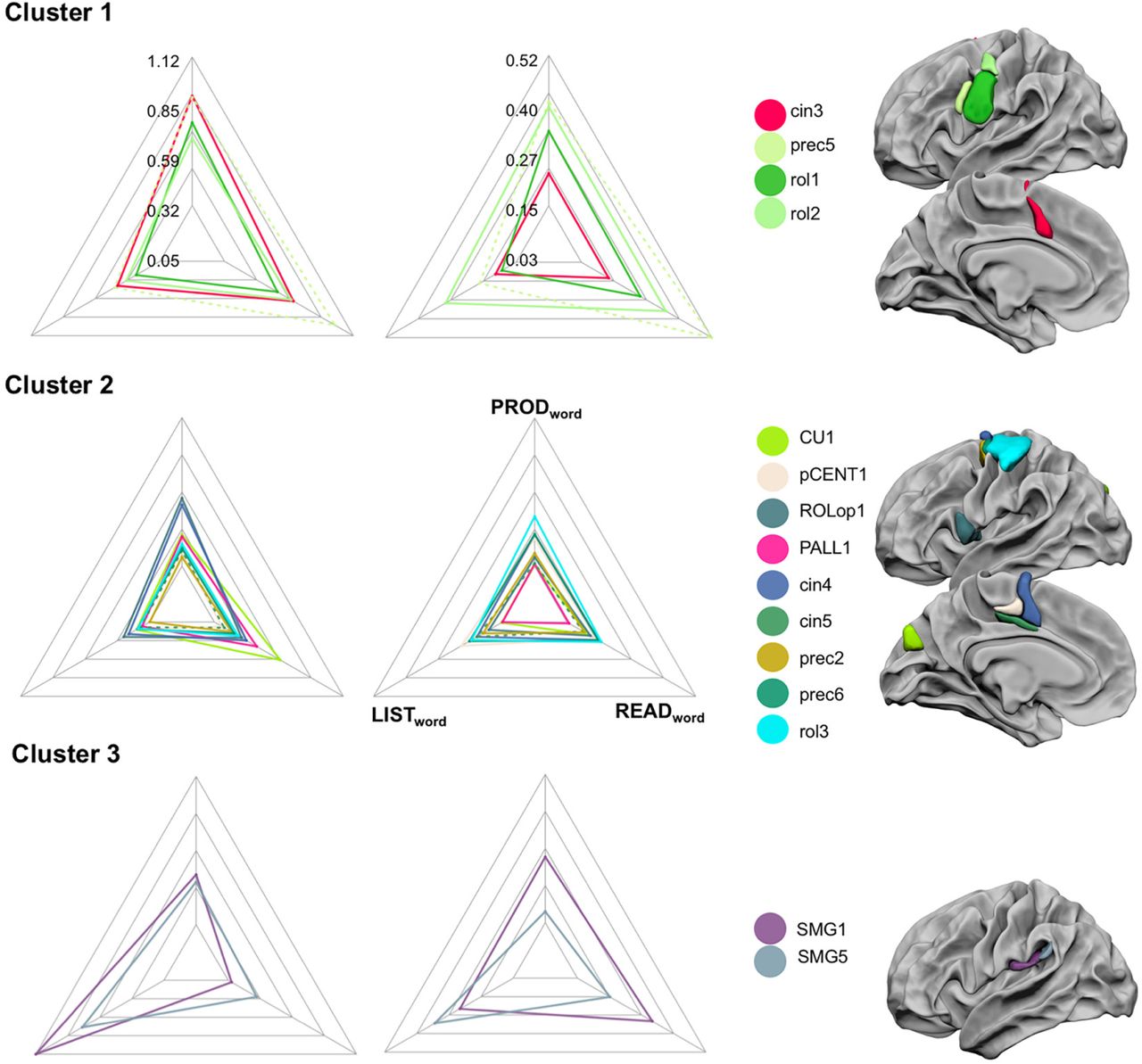
Hierarchical clustering of co-activated and leftward asymmetrical hROIs
The classification obtained by aHCA with the Ward method evidenced 3 clusters of hROIs.
The first cluster included 4 hROIs, all located in the frontal lobe (prec5, rol1, rol2, cing3, Figure 4, Table 2). It was characterized by the high activation strength and strong leftward asymmetry. It was also characterized by a larger activation during PRODWORD and READWORD as compared to LISTWORD (Figure 3 and table 2).

Profile of activation and asymmetry of the hROIs constituting the three left hemisphere clusters. Each row corresponds to a cluster. On a given row, the left radar plot corresponds to activation values, the right radar plot to asymmetry values, the location and color of each hROI constituting the cluster are on the third column. Note that the same scale was applied to the 3 clusters for activation and for asymmetry (provided on the radar plot of cluster 1) and that the same colour-code is applied on the graph and on the hROIs presentation. The values on the top of the radar plots correspond to the BOLD or asymmetry values measured in each hROI during PRODWORD, on the left to those measured during LISTWORD, on the right to those measured during READWORD as shown in the radar plot of asymmetries of cluster 2. Full names of the hROIS corresponding to the abbreviations can be found in table 2.

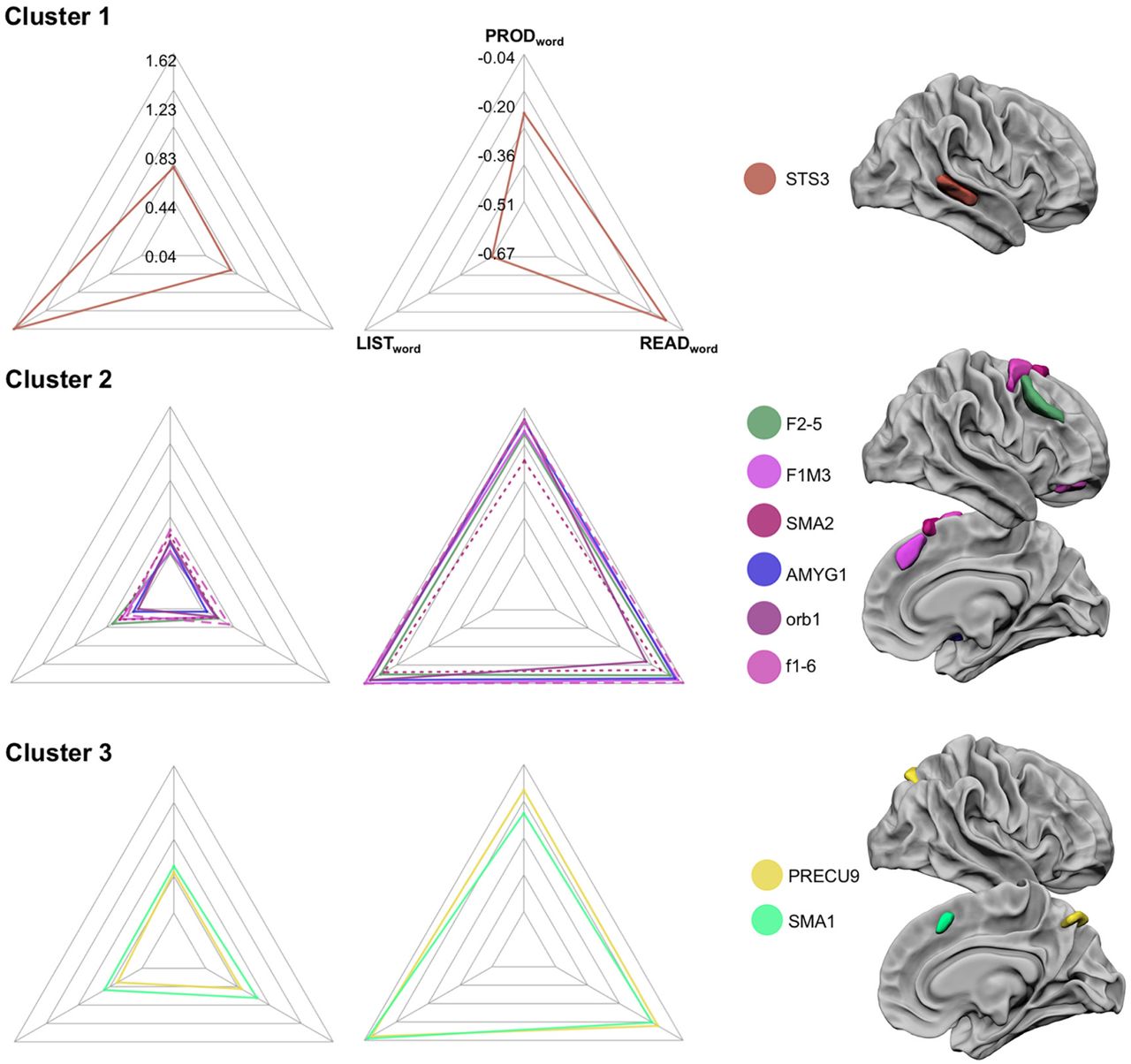
Profile of activation and asymmetry of the hROIs constituting the three right hemisphere clusters, see legend Fig. 3.
MNI coordinates and mean activation in each word list contrasts of the 15 hROIs showing joint left activation and left asymmetry during the 3 tasks organized in clusters. (abb corresponds to the abbreviation of the AICHA hROI name).
The prec5 hROI, located at the junction between the superior frontal sulcus and the precentral sulcus, was particularly activated during READWORD, and post-Hoc analyses revealed significant larger activation than during LISTWORD (p<.001) and PRODWORD (p<0.027) as well as larger leftward asymmetry than during PRODWORD (p<0.01) and LISTWORD (p<.0001, table 3). Within READWORD, Prec5 was more activated and more leftward asymmetrical than adjacent rol1 and rol2 (p<.001).
Mean asymmetry in each word list contrasts of the 15 hROIs showing joint left activation and left asymmetry during the 3 tasks organized in clusters.
The second cluster included 9 frontal hROIs (Figure 3) and was characterized by half lower activation and asymmetries than cluster 1 (Figure 3) and by a low variability of activation and asymmetry across hROIs. It was composed on the lateral surface of 3 adjacent hROIs located at the upper part of the motor and premotor strip (rol3, prec2, prec6), and one frontal area at the rolandic operculum (ROLop1). Three other adjacent hROIs were located in the internal surface of the frontal lobe immediately anterior to the SMA proper (cing4, cing5, ParacLob). Finally, one hROI was located in the dorsal part of the cuneus (CU1) and the last one overlapped the most internal part of the left pallidum (Pall). ROLop1 and cing4 showed higher activation during PRODWORD than during LISTWORD and READWORD (all p<.0001) close to the activation strength of the first cluster. Note that during PRODWORD ROLop1 and cing4 had larger activation than all other hROIs composing the cluster. CU1 had larger activation during READWORD than PRODWORD (p<.0001) and LISTWORD (p<.0001). Note that during READWORD the visual hROI CU1 also showed higher values than the other hROIs of this cluster (p<.0001) even when compared to the PALL (p<0.02) that also showed a high activation during READWORD.
The third cluster included two parietal hROIS located in the supramarginal gyrus (SMG1, SMG5, Figure 3) and was characterized by very strong activation during LISTWORD, as compared to PRODWORD and READWORD (table 2, all post-Hoc p<.0001). For LISTWORD SMG1 was significantly more activated than SMG5 (p<.0001). No significant differences were revealed for asymmetry (table 3).
Reliability and robustness of the identified clusters
The reliability of the 3 clusters identified in the 144 participants, was assessed with the AU p-values provided by the multi-scale bootstrap resampling method. Results showed that the 3 clusters were reliable at levels of 91% for the first cluster, 94% for the second cluster and 97% for the third one.
The classification was quite reliable through the different classification methods. Indeed, the aHCA method with Ward distance compared with (1) the aHCA with the average distance method and (2) the k-Means method led to an adjusted Rand index of 1, indicating a perfect match between these three classifications. On the opposite, the aHCA with Ward distance method compared with the Gaussian Mixture Model, led to an adjusted Rand index of 0.26.
As a whole, the high reliability observed with the bootstrap method together with the consistency of clusterization across the 3 different methods allow for concluding that the classification is robust.
Right hemisphere
Conjunction of regions showing significant rightward activation and asymmetry in the three tasks
The conjunction of significant rightward activation in the three contrasts (intersection of PRODWORD, LISTWORD and READWORD) revealed 65 hROIs (Figure 2, first raw). The conjunction of significant rightward asymmetry evidenced 25 hROIs (Figure 2, second raw) and the joint rightward activation and asymmetry revealed 9 hROIs (Figure 2, third raw): 6 in the frontal cortex, 1 in the temporal cortex, 1 in the parietal cortex and 1 in the amygdala.
Hierarchical clustering of co-activated and asymmetrical right hROIs
The classification by hierarchical clustering with the ward method evidenced 3 clusters of hROIs from the selected set of 9 hROIs.
The first cluster was only made of one region because of its very specific pattern. The STS3 located at the mid-third of the superior temporal sulcus, was actually very strongly activated in LISTWORD (showing the largest right hemisphere activation in the selected hROIs Table 4) as compared to PRODWORD and READWORD (all p<.001). STS3 was also more rightward asymmetrical during LISTWORD than during the two other tasks (Figure 4, table 5, all p<.001).
MNI coordinates and mean activation in each word list contrasts of the 9 hROIs showing joint right activation and right asymmetry during the 3 tasks by cluster.
Mean asymmetry in each word list contrasts of the 9 hROIs showing joint right activation and right asymmetry during the 3 tasks.
The second cluster included 6 hROIs, 5 located in the frontal lobe that had close activation and asymmetry profiles with moderate activation strength and low asymmetries. In terms of activation, F1M1 and F2 had larger activation during READWORD and LISTWORD than during PRODWORD while f6 and orb1 had lower activation during LISTWORD than during the two other tasks (Table 4).
The third cluster, which includes the SMA1 located anteriorly to SMA proper, as well as the PRECU9, was characterized by moderate activations and low asymmetries although asymmetries were larger than those of cluster 2. Both hROIs were slightly less activated and less rightward asymmetrical during LISTWORD as compared to READWORD and PRODWORD (all post-hoc t test p<0.05).
Reliability and robustness of the identified clusters
The reliability of the 3 clusters identified in the 144 participants with the AU p-values provided by the multiscale bootstrap resampling method showed that the 3 clusters were reliable at levels of 100% of reliability for the first cluster (STS3), 56% for the second cluster (SMA2, F1M1, F2, f6, orb1 and AMYG), 55% for the third one (PRECU9 and SMA1). Note that, the second and the third clusters were not strongly different in terms of behaviour (see figure 4), as revealed by the fact that when grouping together, the reliability was at the level of 100%.
The classification was totally reliable using the different classification methods. The aHCA method with Ward distance compared with (1) the aHCA with the average distance method, (2) with the k-Means method, and (3) the Gaussian Mixture model led to an adjusted Rand index of 1, indicating a perfect match between each classification.
Summary of the results
The conjunction analysis of the activations and asymmetries common the PRODWORD, READWORD, and LISTWORD in each hemisphere made it possible to identify the set of areas involved in the processing of lists of familiar words, whatever the modality of the language task. aHCA clustering of the 15 left hROIs selected this way allowed for identifying 3 clusters with different profiles of activation and asymmetry: the first one included motor areas along the central sulcus and premotor preSMA area and exhibited very large activations and larger leftward asymmetries during PRODWORD and READWORD, as compared to LISTWORD; the second had a similar activation profile than the first one but with a lower intensity and no difference in asymmetry across tasks, it included hand motor area and adjacent lateral and internal premotor regions, the Rolandic operculum, and the dorsal part of the precuneus; the third cluster included two SMG hROIs and was much more activated during LISTWORD than during the 2 other tasks. On the right, aHCA clustering of the 9 right hROIs allowed for identifying 3 clusters with different profiles of activation and asymmetry: the first one made of only one region, the STS3, was strongly activated and asymmetrical in LISTWORD as compared to the 2 other conditions. The two others presented close activation and asymmetry profiles across tasks with moderate activation strength and low asymmetries. However, the third one, including SMA1 and PRECU9, had larger activation than those of cluster 2 and mainly included areas located in the superior part of the frontal lobe.
Discussion
Searching for left and right brain areas commonly co-activated and co-asymmetrical during the production, listening and reading tasks allowed for evidencing multimodal language areas involved at the word level. In fact, previous studies have identified brain areas common to reading and speech production (Lindenberg and Scheef, 2007), to reading and listening (Jobard et al., 2007) or to speech production and listening (Pulvermüller et al., 2006), this the first study, to our knowledge, which assesses the neural bases supporting these 3 language tasks. Even if the language material used in this study is a really simple one (lists of months, seasons, days), and the task simple, the statistical power due to the large number of subjects makes it possible to reveal a neural recruitment in each hemisphere common to the three tasks. Actually language processing relies both on linguistic and paralinguistic processing supported by the left and right hemisphere respectively (Gazzaniga, 2000). In the present work the word-lists were composed of over-learnt familiar words purposely chosen to decrease the weight of lexico-semantic while inducing a prosodic processing due to the specific metrics of lists, and the reference task was a motor and attentional task free of any linguistic processing. The linguistic processing involved in the 3 tasks, whatever the modality, thus mainly corresponds to phonological processing, while the paralinguistic aspect was a prosodic processing. To reveal the brain areas specifically involved in each hemisphere, we performed a conjunction of brain activated and asymmetrical areas. Actually, the use of AICHA atlas, which takes into account functionally homotopic regions, makes it possible to reveal leftward asymmetrical language brain regions involved in linguistic and rightward asymmetrical language brain regions involved in paralin-guistic and/or attentional processing. The idea behind the conjunction of activations and asymmetries during the 3 tasks is to be selective enough to present brain areas specifically lateralized during the tasks. Hence, the hierarchical clustering analysis allowed for determining, among the 15 left hROIS and the 9 right hROIS, three left clusters and three right clusters depending on the profiles of hROIS activations and asymmetries during the 3 tasks.
Left activated and asymmetric brain areas involved in word-list processing whatever the task
There does exist a group of hROIs common to the 3 language tasks, with a leftward involvement of specific brain areas. Interestingly, these supra-modal areas spanned the left SMG, whose 2 hROIs were segregated in one cluster, while frontal motor and premotor areas were grouped in 2 clusters presenting 2 specific profiles, as it will be further discussed. These frontal and temporal regions, connected through the arcuate fasciculus tract (Catani et al., 2005), appear to be composed of hROIs involved in phonological processing (Vigneau et al., 2006).
The left perception-action cycle is also involved during reading
The left SMG cluster and the two motor and premotor clusters of the phonological loop also correspond at large to the neural bases of a perception-action cycle for speech sounds commonly recruited during the three language tasks. Such a model, based on the link between speech perception and production, posits that articulatory gestures are the central motor units on which both speech perception and production develop and act according to the motor theory of speech (Liberman and Whalen, 2000). More recently, neurobiological theories of speech perception have proposed a more dynamic and integrative model in which language processing would rely on action-perception circuits distributed across auditory and motor systems (Pulvermüller and Fadiga, 2016, 2010). The fact that clusters 1 and 2 that hosted mostly motor and premotor areas were also activated during LISTWORD, though at a lower intensity and with a lower asymmetry, together with the recruitment of the SMG1 and the SMG5 in the production task, favours the theory supporting an involvement of action-perception circuits whatever the language task (Pulvermüller and Fadiga, 2016, 2010). Actually, it is interesting to observe that READWORD also appears to recruit this action-perception circuit, revealing a significant activation in rol1 as compared to LISTWORD even if it remains less activated than in PRODWORD. The action-perception circuit recruited in the present study for READWORD could reflect the fact that the participants were instructed to engage their attention in the reading of each word (which is supported by the strongest activation in CU1 for the READWORD condition as compared to the 2 other conditions) and covertly articulate the words of the list in a quiet pace. The fact that ROLop1 activation in the present study is similar for READWORD and LISTWORD favours the recruitment of the action-perception circuit reflecting the fact that while reading these over-learnt words, subjects tended to subvocalize them, which is supported by the gradient of significant activation and asymmetry in rol1 (production > reading > listening). Nonetheless, it could also be put forward that ROLop1 activation could implement a simulation of phoneme production according to the model by Wilson & Iacoboni (Wilson and Iacoboni, 2006) which stipulates that the prediction of the acoustic consequences of phoneme production would be compared in the superior temporal cortex with the acoustic analysis of the heard speech sounds. In the present study we cannot disentangle between an actual motor component related to subvocalization from a simulation both being potentially at play depending on the task, i.e., simulation during the listening task and vocalization during the production task as well as a very likely vocalization aspect during the reading task.
More thoroughly, on the action side, results from the hierarchical classification have revealed that motor areas belonging to two different clusters along the left Rolandic and precentral sulcus, were modulated by the mere nature of the task. Regions having the strongest motor coloration were those of cluster 1 located along the Rolandic sulcus and including primary motor areas that showed huge activations in PRODWORD and READWORD. This is in accordance with Penfield’s cortical stimulation studies, which have provided support for the existence of somatotopy within the orofacial region (Penfield and Roberts, 1959). In fact, rol1 and adjacent rol2 belonging to cluster 1 not only matches the area involved in speech production as described by Wilson (Wilson et al., 2004)) but also areas involved in mouth, larynx, tongue, jaw and lips movements, reported by several studies (Brown et al., 2009, 2008; Fox et al., 2001; Grabski et al., 2012; Wilson et al., 2004) (Table 6). More precisely, along the dorsal-to-ventral orientation of the rol1, the representation for speech listening, mouth, larynx, lips, jaw, tongue, larynx clearly resembles the somatotopic organization of speech effectors proposed by Conant and collaborators (Conant et al., 2014). Moreover, the asymmetry in this area was stronger during PRODWORD and READWORD, which is consistent with the fact that these two tasks involve covert articulation or subvocalization. Prec5, tightly joining rol1 and clustered with the above mentioned areas, along with cing3, was characterized by very strong BOLD increase during READWORD as compared to both PRODWORD and LISTWORD, suggesting either motor preparation activity (Dietz et al., 2005) or a role in the retrieval of letter shapes from memory (Liu et al., 2011) or to subvocalization of phonemes while reading a single letter since Longcamp and collaborators have revealed a shift from the left to the right hemisphere in left-handed subjects (Longcamp et al., 2005).
MNI coordinates of speech effectors from the literature.
A second group of hROIs, hierarchically corresponding to increased integration, was more activated by the tasks with a strong motor component even if the strength of activation was moderate as compared to cluster 1. Among these, ROLop1 has been reported to be activated by overt and covert articulation (Heim et al., 2002; Herbster et al., 1997; Price et al., 1996), to be involved in phonological rehearsal (Veroude et al., 2010), in silent recitation of the months of the year (Wildgruber et al., 1996), and whose lesion is associated with articulatory disorders (Tonkonogy and Goodglass, 1981), which is in accordance with its greater involvement in PRODWORD in the current study. Its lower activation as compared to cluster 1 is due to the fact that this region is a secondary motor area, leading to the hypothesis that this secondary motor area, namely, ROLop1 would be part of the perception-action model.
The common activation of the cing3 and cing4 and Pcent1 during the 3 tasks belonging to the second cluster located within the territory of the pre-SMA functional area, with a gradient of activation ranging from more activation for production to less activation for perception, is also in favour of the Motor theory of speech, since pre-SMA activations have been reported to be related to motor planning and imagination of articulation (Indefrey and Levelt, 2004).
In this second cluster one should note that prec2 was also clustered with pre-SMA, 2 regions that were more activated during production and reading and thereby likely to be at the interface between eye fixation and pursuing eye movements, since they overlap the Frontal Eye Field area which plays an important role in the control of visual attention and eye movements (Amiez and Petrides, 2009; Derrfuss et al., 2012). Finally, one should also note that Rol3 which belongs to the same cluster, was commonly activated in the 3 tasks, and was the sole area that did not show any significant difference among tasks. Given its location, the involvement of this region can be directly related to the motor response provided by the subjects at the end of each list of words task (Tzourio-Mazoyer et al., 2015).
Within the large perception-action model one can identify a set of areas that are considered as the neural support of the phonological working memory loop postulated by Baddeley (Baddeley et al., 1998). Actually, dealing with word-list automatically engage working memory processes and that component is common to the 3 language tasks.
On the perception side, one cluster (SMG1) was more activated and asymmetrical in LISTWORD as compared to READWORD and PRODWORD. Interestingly, the SMG1, situated on the posterior part of the planum temporale, corresponding in the literature to the Sylvian-parietal-temporal area or Spt (Buchsbaum et al., 2011; Yue et al., 2018), has been described as a sensory-motor integration area for the vocal tract, whose activity would be correlated with frontal speech production areas even if their precise locations differ according to the studies: the pars opercularis according to Buchsbaum et al, (2001) and Poldrack et al, (Poldrack et al., 1999) and the dorsal part of the pars triangularis of the inferior frontal gyrus according to the meta-analysis by Vigneau et al (2006). Area Spt would be part of the phonological loop described by Baddeley (Baddeley et al., 1998), in which the content of the phonological store can be kept active via articulatory rehearsal (Buchsbaum et al., 2011). More precisely, Spt area has been assumed to have a storage function (Martin, 2005; Smith and Jonides, 1998) and to play an important role in the short-term storage of phonological representations by serving as a phonological buffer (Yue et al., 2018). Together with the Spt activation, on the action side, the Prec5 was found to be co activated and left asymmetrical by the 3 language tasks, and a gradient of activation ranging from more activation for READWORD and PRODWORD and less activation for LISTWORD was revealed. Even if the IFG has not been revealed to be co activated and asymmetrical by the 3 tasks in the current study, due to the bilateral involvement of F3t as revealed by the conjunction of activation (Figure 2, top raw), this premotor area, Prec5, has been proposed to make up a subvocal rehearsal system (Chein and Fiez, 2001) and/or to support executive processes allowing for maintaining the content of verbal working memory (Smith and Jonides, 1998), which is in accordance with the highest activation during READWORD and PRODWORD. Furthermore, a single rTMS intervention targeting either the left SMG or the left pIFG, which are considered as phonological nodes, was already sufficient to disrupt phonological decisions, providing further support that both regions contribute to efficient phonological decisions, and more particularly, subvocal articulation (Hartwigsen et al., 2016).
The Right hemisphere is involved in word-list processing whatever the task
Even if the right hemisphere is known to be involved in language processing, and more particularly in the prosodic dimension of speech (Beaucousin et al., 2007; Sammler et al., 2015), this is the first study that aims at assessing the role of the right hemisphere during both language production, listening and reading lists of words.
The right STS involved in prosodic integration is also activated during PRODWORD and READWORD
The rightward co activation of the STS3 as well as its rightward asymmetry, though more activated in LISTWORD and less activated in READWORD, could be accounted for by a rightward preference for non-linguistic information such as prosody (Belin et al., 2002). More precisely, affective prosody has been revealed to be supported by 2 brain areas in the right temporal lobe, the aSTS and the pSTG (Beaucousin et al., 2007). More particularly, the right STS3, which is located on the internal part of the pSTG, closely matches the pSTG described by Beaucousin and others (2007). This area would be a prosodic integrative area, which would process prosodic cues extracted in the aSTG (Ross, 1981). In the present study, it could be hypothesized that the metrics of lists of the words, resembling a reciting, are processed in this area, which is supported by the greater activation during the listening task. This is also in accordance with the auditory material proposed to the participants, the list being spoken along with the specific prosody of a list. The recruitment of the right AMYG is in line with this hypothesis, since this region has not only been revealed to be involved in voice emotional processing (Adolphs, 2002) but also to lead to deficits in emotional prosodic comprehension when lesioned (Scott et al., 1997). Moreover, even if the methodology used in the present study does not permit to assess the functional connectivity within brain networks, it has been reported that during speech, words present non-verbal prosodic information which is intertwined with the verbal information (Kotz and Paulmann, 2007; Pell and Kotz, 2011). Furthermore, prosodic and verbal cues in speech differ in their spectrotemporal properties: prosodic cues consisting of slow spectral changes (over more than 200ms) and phonemic cues consisting of fast spectral changes (less than 50ms). The right hemisphere has been suggested to be more sensitive to fine spectral details over relatively long time scales and the left hemisphere more sensitive to brief spectral changes (Boemio et al., 2005; Poeppel et al., 2008, 2008; Zatorre and Belin, 2001). The existence of a link between the activity of the right ST3, which is a prosodic integrative area with that of the left SMG5, thus reflecting the intertwining between prosodic and phonemic information could be hypothesized. This hypothesis is supported by a recent fMRI/ERP study which has revealed activity in the left SMG together with the central sulcus area during the phonological processing of ambiguous speech syllables, while attention to the spectral (prosodic) properties of the same stimuli leads to activity in the right STS (Liebenthal et al., 2016).
Right visuospatial and attentional areas
Brain areas constituting clusters 2 and 3 presented low difference between the 3 tasks, either in activation or asymmetry, meaning that they were more related to the attentional processes share in common by of the tasks. In fact, in the 3 conditions, once the task was completed, subjects had to detect the transformation of a centrally displayed cross into a square and they were asked to press a button. One should note that even if it is a low reference task, it elicits specific motor and visual attentional processes since there is no reference condition. Among these areas, the SMA2 and F1M3 overlapping location of the supplementary frontal eye fields together with the posterior part of the middle frontal gyrus correspond to the dorsal frontal attentional network (Corbetta and Shulman, 2011) while the Precuneus could be related to mental imagery triggered by the scramble version of the picture or to episodic memory encoding in reference to the list of the days and months (Cavanna and Trimble, 2006).
General conclusion and perspectives
The present study, based on the fMRI analysis of 3 language tasks performed by 144 healthy adult right-handers reveals, for the first time, the existence of hetero-modal areas common to production, comprehension and reading at the word level. Interestingly, leftward frontal and precentral areas together with temporo-parietal areas have been revealed to be commonly activated and asymmetrical, leading to the hypothesis of the involvement of phonological action perception circuits such as the phonological working memory loop, in which articulatory gestures are the central motor units on which word perception, production and reading would develop and act according to the motor theory of speech (Liberman and Whalen, 2000). On the right hemisphere, the common activation and asymmetry of the STS3, which is a prosodic integrative area, during the 3 tasks, could reflect the processing of fine spectral details over time. Future research based on the analysis of intrinsic resting-state connectivity would make it possible to assess the existence of brain networks during reading, listening and production tasks.
General conclusion and perspectives
The present study, based on the fMRI analysis of 3 language tasks performed by 144 healthy adult right-handers reveals, for the first time, the existence of hetero-modal areas common to production, comprehension and reading at the word level. Interestingly, leftward frontal and precentral areas together with temporo-parietal areas have been revealed to be commonly activated and asymmetrical, leading to the hypothesis of the involvement of phonological action perception circuits such as the phonological working memory loop, in which articulatory gestures are the central motor units on which word perception, production and reading would develop and act according to the motor theory of speech (Liberman and Whalen, 2000). On the right hemisphere, the common activation and asymmetry of the STS3, which is a prosodic integrative area, during the 3 tasks, could reflect the processing of fine spectral details over time. Future research based on the analysis of intrinsic resting-state connectivity would make it possible to assess the existence of brain networks during reading, listening and production tasks.
Funding Sources
This work was supported by a grant from the FLAG-ERA Human Brain Project 2015 (ANR-15-HBPR-0001–03-MULTI-LATERAL) to NTM and by a research contract from CEA to LL.
Supplementary materials
Regional variations in LIST activations and asymmetry maps for each task
Production (Fig. S1)
Regions of the AICHA atlas significantly activated or asymmetrical during word-lists production. The hROIs are presented on the white matter surface of the BIL&GIN template, regions significantly activated on the lateral surface are presented on the first raw and those of the internal surface on the second row, left hemisphere is on the left, and right hemisphere on the right. Selection of hROIs considered as activated or asymmetrical per hemisphere was done according to Bonferroni (p corrected <.00027, 184 hROIs tested).
Activation
During the production of lists of words, 111 leftward hROIs and 127 rightward hROIs were activated, leading to the recruitment of a bilateral network encompassing the frontal cortices from the SMA up to the middle cingulate gyrus on the one hand, and along the precentral gyrus up to the IFG (pars opercularis, triangularis and orbitaris) on the other hand. Bilateral hROIs activations were also revealed in the superior temporal cortices, the parietal cortices (postcentral gyrus), the occipital cortices and the insula.
Asymmetry
44 leftward asymmetric hROIs encompassing the SMA, the precentral gyrus and the posterior temporal gyrus were revealed. 92 rightward hROIs were distributed in the SMA, the superior frontal cortex, the pars orbitaris of the IFG, the temporal gyrus, the precuneus and cuneus, the SMG and in the middle occipital gyrus.
Listening (Fig. S2)
Activation
During the perception of lists of words, 90 leftward and 96 rightward hROIs were recruited leading to bilateral activations in the temporal cortices (primary and secondary areas), the frontal cortices from the SMA, along the precentral gyrus up to the IFG (pars opercularis, triangularis and orbitaris).
Asymmetry
62 leftward hROIs encompassing the SMA, the middle frontal cortex, the precentral cortex, the superior and middle temporal cortex down to the parahippocampal region, the inferior parietal cortex and the occipital cortex were activated. 62 rightward hROIs encompassing the SMA, the superior frontal cortex, the pars triangularis and opercularis of the IFG, down to the middle cingulate, the middle temporal cortex, the inferior parietal cortex and the thalamus were activated.

Regions of the AICHA atlas significantly activated or asymmetrical during word-lists listening. The hROIs are presented on the white matter surface of the BIL&GIN template, regions significantly activated on the lateral surface are presented on the first raw and those of the internal surface on the second row, left hemisphere is on the left, and right hemisphere on the right. Selection of hROIs considered as activated or asymmetrical per hemisphere was done according to Bonferroni (p corrected <.00027, 184 hROIs tested).
Reading (Fig. S3)
Activation
During the reading of lists of words, 110 leftward and 111 rightward hROIs were recruited, leading to bilateral activations in the occipital cortices, the frontal cortices from the SMA, along the precentral gyrus up to the IFG and the parietal cortices (postcentral gyrus).
Asymmetry
58 leftward hROIs encompassing the SMA, the precentral gyrus, the superior temporal gyrus, the SMG, the superior and middle occipital gyrus as well as the thalamus and the putamen were activated. 76 rightward hROIs encompassing the SMA, the middle frontal cortex down to the middle cingulate cortex, the superior and inferior temporal cortex as well as the parahippocampal region, the inferior parietal cortex and the middle occipital cortex were activated.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Regions of the AICHA atlas significantly activated or asymmetrical during word-lists reading. The hROIs are presented on the white matter surface of the BIL&GIN template, regions significantly activated on the lateral surface are presented on the first raw and those of the internal surface on the second row, left hemisphere is on the left, and right hemisphere on the right. Selection of hROIs considered as activated or asymmetrical per hemisphere was done according to Bonferroni (p corrected <.00027, 184 hROIs tested).
Conjunction in LIST activations and asymmetry maps for each task
Word-list production: joint activation and asymmetry (bottom Fig.S1)
Joint left activation and left asymmetry revealed activation in 31 hROIs distributed along the precentral sulcus down to the Rolandic sulcus as well as in the internal surface encompassing the mid cingulum and the cingulate.
Joint right activation and right asymmetry revealed 68 rightward hROIs activation in the inferior and lateral occipital lobe extending to the SMG and along the temporal gyrus up to the pole. The SMA, the middle frontal cortex, the middle orbitaris cortex as well as the anterior insula were also recruited.
Word-list listening: joint activation and asymmetry (bottom Fig.S2)
Joint left activation and left asymmetry revealed 35 hROIs encompassing the Rolandic sulcus, the Rolandic opercularis gyrus and the precentral gyrus, extending to the posterior and anterior insula as well as the SMG. Some peaks of activation were also revealed in the superior and middle orbitaris gyrus and the inferior temporal gyrus.
Joint right activation and asymmetry revealed 36 hROIs encompassing the superior temporal sulcus, the middle temporal gyrus and the temporal pole, the SMG and the AG and the precuneus. Some peaks of activation were also revealed from the SMA, along the precentral sulcus, the mid frontal gyrus and the pars triangularis of the inferior frontal gyrus.
Word-list reading: joint activation and asymmetry (bottom Fig.S3)
Joint left activation and left asymmetry revealed 42 hROIs encompassing the inferior, middle and lateral occipital gyri, the fusiform gyrus, the parieto-occipital sulcus, the cuneus and the superior parietal gyrus as well as the superior temporal sulcus and the SMG. hROIs were also situated along the precentral sulcus and the Rolando sulcus down to the Rolandic operculum.
Joint right activation and asymmetry revealed 46 hROIs encompassing the superior, middle and lateral occipital gyrus, the lingual gyrus and the precuneus. Peaks of activation were also revealed in the inferior parietal gyrus, the SMG, the superior temporal sulcus and the temporal pole. In the frontal lobe, hROIs encompassing the SMA, the superior frontal gyrus, the middle frontal gyrus, the inferior orbitaris frontal gyrus, as well as the anterior insular gyrus were revealed.
Acknowledgements
The authors would like to thank Sandrine Cremona for her help concerning artwork.
References




