

Abstract
Making perceptual decisions to categorize unknown sensory stimuli is a fundamental cognitive function, known as category learning. The posterior parietal cortex (PPC), although has been intensively studied for its role in decision-making and other cognitive functions, its causal link with behavior remains controversial. Here we combine in vivo two-photon imaging, circuit manipulation and auditory psychophysics behavior in mice to probe the role of PPC and its connectivity with sensory cortex in decision-making during category learning. We show that PPC neuronal populations exhibit coding dynamics characteristic of category learning, showing representations for both new sensory stimuli and prior learned categories. Circuit-specific perturbations of PPC and its projections to auditory cortex impaired decision performance specifically for categorizing new auditory stimuli. These data reveal a dynamic and causal role of the parietal-auditory circuit in decision-making, integrating prior knowledge to guide categorical decisions on new sensory stimuli.
Introduction
The ability of humans and other animals to classify unknown items into behaviorally relevant categories or concepts, known as category learning, is essential for survival and fundamental to higher cognition (1–5). For example, to decide a newly encountered animal as a predator or prey, or an unknown plant as nutrients or poisons, the subject needs to compare the perceptual properties of the unfamiliar items with prior experiences. Central to this process is making perceptual judgements on novel sensory stimuli based on their perceptual relationship to prior learned prototypes or exemplars, an essential step in category learning. The underlying causal neural circuit causal mechanism remains poorly understood. The posterior parietal cortex (PPC) has been widely reported to show decision and categorization-related activity both in primates (6–12) and in rodents (13–17), and has also been implicated in many other cognitive functions (8,14,18–22). Therefore, PPC and its connectivity with related regions are likely to be the key brain system in category learning. However, recent studies reported rather controversial results regarding the causal role of PPC in choice behavior. In macaques, inactivation of PPC did not have significant effect on behavioral performance in the random-dot visual motion decision task (23) or a self-motion perceptual decision task (24). In rodents, silencing PPC neither had significant behavioral effect in auditory or tactile decision tasks (17, 25, 26). But silencing or disruption of PPC activity impaired visually guided choice behavior in rodents (14, 17, 27, 28). Thus, it has become controversial whether PPC is functionally significant for general decisionmaking processes, or it is only required specifically in visual-related functions in rodents.
Most of these studies, however, focused on the PPC activity after animals were well-trained by the same set of sensory stimuli also being used for assessing performance following perturbation, whereas in the real world, animals make decisions upon unfamiliar sensory stimuli based on their relationship to prior learned exemplars or prototypes, a learning capability to survive dynamic and changing environments (1–5). We thus sought to determine the role of PPC in categorical decision-making upon new sensory stimuli that were not used for prior training. Combining mouse auditory psychophysics behavior with in vivo two-photon calcium imaging and circuit manipulations, we demonstrate that PPC and its connectivity with the auditory cortex are dynamically and causally involved in categorical decision-making on new sensory stimuli, providing a causal circuit mechanism for category learning.
Auditory category learning in head-fixed mice
We trained head-fixed mice to perform a two-alternative-forced-choice (2AFC) auditory discrimination task using two exemplar tones (training stimuli, 2 octaves apart, 8 kHz and 32 kHz; Fig. 1A). Mice report their perceptual decisions on tone stimuli as “low” or “high” frequency by licking the left or right lick-port (Fig. 1A; see fig. S1 and movie S1). We then introduced new “test” tones in randomly interleaved trials (30%) to examine whether mice can make categorical judgements on new sensory stimuli based on prior trainings (Fig. 1A). Remarkably, mice exhibit sharp categorical decisions for the test stimuli (Fig. 1, B and C), indicated by the peak discriminability (the slope value at the categorization boundary) significantly greater than that for an otherwise linear relation between choice and tone frequency (Fig. 1, C and D). Mice’s subjective categorization boundaries (points of subjective equality, PSE) are distributed in close proximity of experimentally defined category boundary (16 kHz; Fig. 1E; also see fig. S1E). These data indicate that mice are able to make categorical decisions to classify new tone stimuli as “high” or “low” based on previous trainings, a key process in category learning (29).
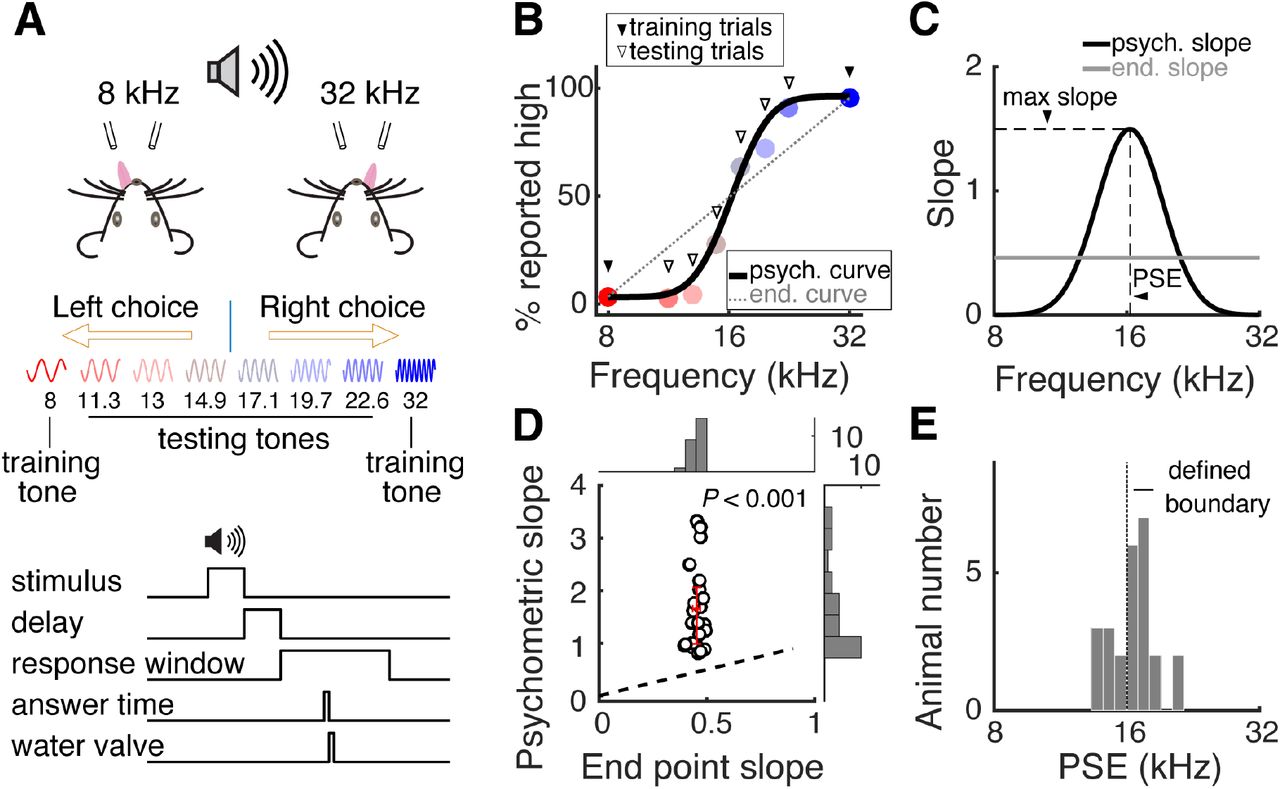
(A) Schematic showing the configuration of behavioral paradigm and trial structure. (B) Psychometric function from an example behavioral session where the test stimuli were first introduced. (C) Slopes of the psychometric function and hypothetical linear relation between stimulus and behavioral choice shown in (B) The peak of the slope of the psychometric function (black) around the point of subjective equality (PSE) indicates perceptual categorization. The slope of the linear function (grey) shows a constant value. (D) Scatter plot of group data comparing peak psychometric slopes (1.66 ± 0.16) and linear relation slopes (0.45 ± 0.01) within each animal (P < 0.0001, n = 25 mice). (E) Histogram showing distribution of the PSE of 25 animals from the first session introducing test stimuli (PSE = 16.5 ± 0.3 kHz). Red error bars indicate median and interquartile range.
PPC neurons exhibit characteristic dynamics of category learning
Although the dynamics of PPC activity has been recently studied (13, 14, 16, 21, 30), the precise relationship between PPC activity and specific learning process is still unclear. Here we investigate the relationship between PPC activity dynamics and perceptual category learning, using chronic in vivo two-photon calcium imaging from PPC neurons across different sessions/ days. Genetically encoded calcium indicator GCaMP6s (31) was expressed in PPC of left hemisphere using AAV vectors to indicate the intracellular calcium dynamics, followed by implantation of a chronic imaging window (32). The same population of layer 2/3 neurons were imaged during task performance repeatedly across days starting from the session when the test stimuli were first introduced (Fig. 2A; fig. 2; also see methods in supplementary materials). Consistent with previous studies (14, 16, 22, 27), we found that PPC neurons show complex response selectivity to multiple behavioral variables (Fig. 2B; fig. 3; table S1).

(A) Schematic and images showing in vivo two-photon imaging from PPC populations using GCaMP6s. The same imaging fields were repeatedly imaged across days. (B) Example Ca2+ signal traces of PPC neurons showing high signal to noise ratio with diverse selectivity and rich dynamics. (C) Ca2+ signal of an example PPC neuron imaged during behavioral sessions on consecutive days. Upper, color indicates % ΔF/F0. Blocks of trials presenting training and test stimuli of low and high frequency categories were segregated. Trials within each block were aligned by stimulus onset time and sorted by time of first answer lick. Lower, mean Ca2+ signal traces averaged from blocks of trials in the upper panel. Color corresponding to tone frequencies. (D) Mean response amplitudes in 1 s window after stimulus onset averaged from different blocks shown in (C). Error bars, s.e.m. (E) Selectivity index (SI) between training and test stimuli across time based on ROC analysis using the neuronal responses as shown in c, for all neurons in an example imaging field. Positive values of SI represent preference to test stimuli and negative value for training stimuli. (F) Histogram showing distributions of SI values (mean value in 1 s window after stimulus onset) for training vs. test stimuli from the total of 757 neurons across two consecutive days. Significant preference to test stimuli on day 1 (P < 10-23, Wilcoxon signed rank test), but to training stimuli on day 2 (P < 10−6, Wilcoxon signed rank test). (G) Averaged area under ROC curve, discriminating training and test stimuli, across consecutive days for neurons with significant preference to test stimuli on day 1. Error bars, s.e.m. (H) Color raster plot showing index of selectivity between high and low frequency categories across trial time, for all neurons as in (F) across consecutive days. Neurons are sorted according to SI values on day 1. (I) Scatter plot of the mean SI values in the 1 s trial time window after stimulus onset as in (H) r, correlation coefficient. Slope, slope of the linear regression line (red). (J) Decoding accuracy across trial time for high and low training tones by population activity of simultaneously imaged neurons in each imaging field using a linear classifier. The classifier was trained using neuronal activity in 80% of training trials on day 1 to decode stimulus categories on consecutive 3 days. Insert, mean decoding accuracies in a 1 sec time window following stimulus onset. (K) Schematic showing a conceptual model for differential dynamic representations in PPC reflecting category learning.

(A) Schematic and histology image showing the locations of muscimol or saline injection in PPC. Red color in the histology image is the fluorescence of co-injected red beads to mark the injection sites. (B) Schematic and histology image showing the locations of photo stimulation and viral expression of ChR2-mCherry in GABAergic neurons in bilateral PPC of Vgat-Cre mice. Red fluorescence represents mCherry expression. (C), Schematic showing experimental schedules for PPC inactivation and control experiments on the first day introducing test stimuli. Symbols represent the 4 types of experiments as indicated in (A) and (B). Frequencies of tone stimulation at different stages were indicated. (D) Psychometric functions (upper) and corresponding slopes (lower) from an example muscimol injection session and control session as indicated in (A) and (C). Error bars, 95% confidence interval. See fig. S9 for individual cases. (E) Similar as in (D) for an photoinhibition and control sessions as indicated in (B) and (C). See fig. S10 for individual cases. (F) Summary of grouped data of peak psychometric slopes from experiments indicated in (C). Muscimol, n = 13, vs. saline, n = 14, P < 0.05; photoinhibition, n = 7, vs. mask only, n = 6, P < 0.05. Error bars, s.e.m. (G) Summary of PSE for experiments indicated in (C) (muscimol vs. saline, P = 0.68; photoinhibition with mask vs. mask only, P = 0.73; Error bars, s.e.m.). (H) Summary of mice’s performance (percent correct) for training stimuli, compared between muscimol injection and control experiments. (I) Similar as in (H) for photoinhibition vs. control experiments. Error bars, s.e.m. Paired ŕ-test. (J) Experimental schedules, similar as in (C), for PPC inactivation on the third day after introducing test stimuli. (K) Similar plot as in e for photoinhibition experiments indicated in (J). See fig. S11 for individual cases. (L) and (M), Summary of grouped data of peak psychometric slopes for experiments indicated in (K).
Here we ask whether the PPC neuronal dynamics exhibits characteristics of category learning. With the progress of learning, new sensory stimuli are classified into existing categories, such that the perceptual distinction between new sensory stimuli and exemplars of the same category tend to diminish. We thus focused on whether PPC neuronal activity could distinguish the test stimuli (new sensory information) and the training stimuli (prior knowledge) of the same category, and how such distinction changes with learning. The high performance of perceptual categorization in mice for the test stimuli on day 1 (the first session introducing test stimuli; Fig. 1) suggests that significant category learning likely occurred rapidly. Indeed, we found rapid changes in PPC neuronal responses to test stimuli across the first two sessions (days) following introducing test stimuli. On day 1, a significant fraction of neurons responded more strongly to test stimuli than to training stimuli, however, such difference was markedly reduced on day 2 (Fig. 2, C and D; also see fig. S4).
To quantify the distinction in neuronal responses between test stimuli and training stimuli of the same category, we use receiver operator characteristic (ROC) analysis to define a selectivity index (SI), with negative SI indicating stronger preference to training trials, and positive SI, stronger preference to testing trials (see methods in supplementary materials) (33). For learning, the activity in the entire trial is likely to be important, we therefore computed the SI of each neuron in an imaging field for every imaging frame across the time course of the trial (Fig. 2E). We found that on day 1, when the test stimuli were first introduced, more neurons significantly distinguish test stimuli and training stimuli, with stronger preference to test stimuli. However, on day 2 and day 3, the preference to test stimuli were largely reduced. This trend is also evident over the entire neuronal population (757 neurons from 6 mice), where the population distribution of SIs also shifted from significantly positive values on day 1 (P<0.001, signed Wilcoxon rank sum test) toward zero on day 2 (Fig. 2F). For neurons showing significant preference to test stimuli on day 1, the neuronal discrimination (measured by the area under ROC curve) between test stimuli and training stimuli of the same category also significantly decreased in day 2 and 3 (Fig. 2G), indicating that the distinction between the test stimuli and training stimuli of the same category was largely reduced. These results indicate that in parallel to perceptual categorization, PPC neuronal representations exhibit the characteristic dynamics of category learning, incorporating new sensory stimuli into existing categories.
During learning, the circuits not only need to dynamically integrate new information but also need to maintain a stable representation of prior knowledge. We thus ask whether the representation of established “high” and “low” categories in PPC population is relatively stable. First, we found that the number of neurons selective to training stimuli was almost unchanged (fig. S5E; 21% in day 1 vs. 20% in day 2). Second, using ROC analysis, we computed the selectivity index of each neuron for “high” vs. “low” frequency categories for all the neurons (757 neurons) over the time course of the behavioral trial across consecutive days (Fig. 2H; see methods in supplementary materials). We sort all the neurons according to their SI values on day 1. We found that across the entire population and trial time course, neuronal selectivity for “high” and “low” categories is highly preserved across days, as indicated by the resemblance of the profiles of the SIs between the two consecutive days (Fig. 2H; Pearson’s r = 0.83). The mean SI values within the time epoch of stimulus and choice (1 s time window after stimulus onset) are also strongly correlated between two consecutive days (Fig. 2I; r = 0.91). Thus, the neuronal selectivity for established stimulus categories is stable across days. Third, we asked whether the population coding for the established categories was also stable. We used a linear classifier (34) to examine the population decoding accuracy of simultaneously imaged neurons in each imaging field for categorical discrimination over the trial time. We used the population activity in 80% of training trials on day 1 as training set to predict either the rest of training trials in day 1, or the training trials in day 2 and day 3 respectively (Fig. 2J). We found that the decoding accuracies around the decision time (first lick) were not significantly different for day 1, day 2, and day 3 (Fig. 2J), indicating that the population coding for the stimulus categories was stable across days over category learning. Taken together, these results show that PPC neurons exhibit characteristic dynamics of category learning, adaptively classifying new sensory stimuli into existing categories, while maintaining a stable representation for prior learned categories (Fig. 2K).
PPC activity is necessary for perceptual decision-making during category learning
Next, we sought to determine whether PPC activity is important in the decision process during category learning. Our imaging and behavioral data suggest that the learning occurred primarily when the test stimuli were initially introduced in day 1, therefore, it is likely that PPC activity may play a critical role during this period. To test this, we reversibly silenced PPC by bilateral microinjection of muscimol, a selective GABAA receptor agonist, when the new test stimuli were first introduced (Fig. 3, A and C). In contrast to previous reports in rodents (17, 22, 26, 28) and primates (23, 24), we found that mice’s decisions on newly introduced test stimuli were markedly impaired, comparing to control mice microinjected with saline also on day 1. The impairment is measured as a significant reduction in the peak discriminability (slope at the category boundary; saline, 1.46 ± 0.16; muscimol, 0.97 ± 0.09; P < 0.05, Wilcoxon rank sum test; Fig. 3, D and F).
Since muscimol inactivation does not have the temporal resolution to distinguish whether it was during training trials or during testing trials that silencing PPC led to impaired categorization, we employed optogenetics to inactivate PPC in a more temporally controlled fashion (25). An adeno-associated virus (AAV) encoding Cre-inducible channelrhodopsin 2 (ChR2) (35) was injected bilaterally in PPC of VGAT-Cre mice (36) to express ChR2 in GABAergic inhibitory neurons. Glass optical windows were implanted above PPC of both hemispheres following virus injection. Collimated blue laser beams were used to illuminate the glass window to activate ChR2 in GABAergic neurons to inhibit the illuminated cortical areas (Fig. 3, B and C; also see Movie S1) (25). During task performance, photoinhibition was applied in all testing trials, but only in 10% of training trials for comparison, leaving majority (90%) of training trials unaffected. We found that this manipulation also significantly reduced mice’s categorization performance for the new tone stimuli (control, 2.43 ± 0.46; photoinhibition, 1.17 ± 0.37; P < 0.05, Wilcoxon rank sum test; Fig. 3, E and F), indicating that PPC activity during testing trials rather than during training trials is required for categorical decisions on the new test stimuli. Moreover, we found that when photoinhibition of PPC was applied in 50% of the testing trials, mice’s categorization performance was not significantly different in photostimulated trials comparing to that in control trials (fig. 7), suggesting that PPC activity in partial of the testing trials on day 1 was sufficient for category learning of the newly introduced tone stimuli.
These results, together with our imaging data (Fig. 2), point to the possibility that PPC activity is specifically required only during category learning for new stimuli, but not after learning. Consistent with this idea, for the training stimuli, neither muscimol silencing (Fig. 3H), nor photoinhibition (Fig. 3I) influenced mice’s performance (also see fig. 8). To further test this possibility, we performed bilateral PPC perturbation in a separate cohort of mice after the test stimuli had been well-experienced following continued training for two additional sessions (>1000 trials; Fig. 3J). We found that when the test stimuli were well-experienced, inactivation of PPC did not significantly influence mice’s categorization performance (Fig. 3, K to M), agreeing with previous studies in rodents and non-human primates assessing the effect of PPC inactivation on choice behavior, where the stimuli were likely also well-learned (17, 23, 25, 26). Taken together, these data indicate that PPC is causally involved in categorical decisions specifically on new sensory stimuli during category learning, but not after learning when the stimuli have been already incorporated in to existing categories.
PPC activity counterbalances biases from recent trial history
Decision-making in humans and animals is often biased by recent behavioral history, which usually produce a negative impact on performance in complex and unpredictable settings (21, 22, 37–41). It has been reported that PPC activity reflected both accumulated behavioral history (15, 16, 21, 22) and systematic bias from long-term training history (8). Meanwhile, our imaging data, as well as previous studies (6, 11, 15, 24), show that even after learning, PPC neurons continue to encode stimulus category and choice information. To understand how the accumulated prior knowledge encoded in PPC may influence the bias from recent behavioral history, we examined the effect of PPC inactivation on trial-by-trial history bias during the auditory categorization task. We quantified the influence of the preceding training trials on choices for testing stimuli by comparing mice’s performance on test stimuli when the stimulus in the immediate preceding training trial was of the same vs. different category as the current test stimuli. Under control condition, preceding training trials did not significantly bias mice’s performance on the testing trials (Fig. 4B, 77.3 ± 4.6% for same category vs. 74.3 ± 3.2% for different category, P = 0.32), consistent with a previous study, in cases where decisions did not depend on short-term memory (22). However, when PPC was inactivated by either muscimol or photoinhibition, preceding training trials significantly biased mice’s choices on test stimuli towards the preceding categories (Fig. 4A, muscimol, same 71.8 ± 3.4% vs. different 63.5 ± 3.7%, P = 0.017; photoinhibition, same 70.3 ± 2.4% vs. different 55.9±3.2%, P = 0.003.) Thus, whereas mice’s decisions were minimally influenced by preceding events when PPC was intact, inactivation of PPC led to a significant bias by short-term behavioral history (Fig. 4, C and D). Moreover, when the newly introduced test stimuli became well-experienced after continued training, inactivation of PPC no longer resulted in significant bias by preceding training trials (fig. S12, E and F). These results indicate that PPC activity counterbalances the short-term history bias when categorizing novel sensory stimuli.

(A) Comparison of behavioral performance on testing trials when the stimuli in preceding training trial was of different categories (diff. prev.) or the same category (same prev.) as the current trial. Mice’s choices were significantly biased by the previous trial when PPC was silenced by muscimol (P = 0.017, n = 13) or by photoinhibition (P = 0.003, n = 7). (B) Same plot as in (A). Mice’s choices were not significantly biased by previous trials under control conditions (saline, P = 0.14, n = 14; light mask, P = 0.32, n = 6). (C) Summary of changes in performance attributable to previous trials as in (A) and (B). Data from a separate group of control mice without any manipulation were also included. *, P < 0.05; **, P < 0.01. Two sample t-test. Error bars, s.e.m. (D) Schematic showing the bias effect on categorical decisions by the previous trials when PPC was silenced. (E) Illustration of linear regression models to simultaneously quantify the effects of PPC silencing on categorization slopes and short-term history bias. (F) to (H), Likelihood ratio test comparing different regression models using data from the control group for models with or without previous trial choices (F), using all data from both control and PPC silencing groups for models with or without the interaction term between sensory stimuli and PPC silencing (G), and using data from both groups for models with or without the interaction term between PPC silencing and previous choices (H).
To simultaneously examine PPC’s contributions to category learning (Fig. 3) and to short-term history bias (Fig. 4, A and C), we modeled animals’ performance in control and silencing conditions using a series of logistic regression models (Generalized Linear Mixed-Model, GLMM; Fig. 4E; also see methods in supplementary materials) (39). Our models confirmed the lack of history bias effect in control data, in that including of previous choice as a regressor term did not significantly improved the model prediction of the behavioral data (Fig. 4F). The models also confirmed the effect of PPC inactivation on changing the psychometric slopes, in that including of PPC silencing as a regressor significantly improved the model prediction (Fig. 4G; also see fig. S13, A to D). Interestingly, when including an interaction term between previous choice and the silencing condition as a regressor, the model predicted animals’ behavior significantly better comparing to the model that only quantified the effect of silencing on the psychometric slope changes (Fig. 4H; see fig. S14, A to C), indicating that the effect of PPC silencing on short-term history bias is in addition to its effect on categorization slopes. Thus, PPC causally contributes to the decision process in category learning by both influencing categorization slopes and counterbalancing short-term history bias when categorizing novel stimuli.
PPC to auditory cortex projections are important for categorical auditory decisions
PPC neurons consist of diverse populations interconnected with many brain regions (42), and are involved in many behavioral functions (6, 7, 9, 14, 18, 43). It is likely that for a given task, specific circuits interconnecting PPC and task-specific brain regions are critically involved. We thus further examined the circuit-level specificity of the causal role of PPC neurons in auditory categorical decision-making by manipulating PPC to auditory cortex projections using chemogenetics. We first determined the projection patterns between PPC and the auditory cortex by expressing green fluorescent proteins (GFP) in PPC, and red fluorescent proteins (tdTomato) in the primary auditory cortex (AUDp), using AAV vectors. We found that PPC receives projections from AUDp while sending projections to the dorsal portion of auditory cortex (AUDd; Fig. 5A), implying an interconnected circuit between PPC and the auditory cortex (Fig. 5B). We next expressed hM4Di, a designer receptor exclusively activated by designer drug (DREADD), in PPC of both hemispheres using an AAV vector (AAV-Syn-hM4Di). On day 1 (test tones first introduced), we injected Clozapine-N-oxide (CNO) to bilateral AUDd to silence the projections from PPC prior the behavioral session (Fig. 5C). As control, in a separate group of mice we injected AAV-Syn-mCherry in PPC, and then injected CNO to AUDd during task performance on day 1. We found that comparing to the control group, silencing PPC to AUDd projections impaired mice’s categorization performance for the new tone stimuli, as indicated by a significant reduction in the slopes of psychometric functions (Fig. 5D). Quantification using GLMM test also shows a significantly better prediction of the model with the term of projection silencing than without the term (Fig. 5E). These results indicate that the specific projections from PPC to secondary auditory cortex play a critical role in the auditory categorical decisions on new sensory stimuli.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Histology images showing the projection patterns between PPC and the auditory cortex. Left, red fluorescence indicates the site of injection of AAV-syn-mCherry in the primary auditory cortex. Green fluorescence in the dorsal auditory cortex indicates axons projected from PPC. Insert, showing site of injection of AAV-syn-GFP in PPC. Right, red fluorescence indicates axons projected from the primary auditory cortex in PPC. (B) Schematic showing the projection patterns between PPC and the auditory cortex. (C) Schematic showing chemogenetic silencing of the projections from PPC to the auditory cortex. AAV-syn-hM4D-mCherry (experimental group) or AAV-syn-mCherry (control group) were injected in bilateral PPC. CNO was microinjected in bilateral auditory cortex prior the first session introducing test tone stimuli. (D) Summary of the effect of silencing PPC to auditory cortex projections on psychometric slopes. P < 0.05, Wilcoxon rank sum test, n (control) = 7, n (silencing) = 8. Error bars, s.e.m. (E) Likelihood ratio test comparing regression models with or without projection silencing as a regressor.
The neural basis for how organisms integrate novel sensory information with prior knowledge to assign behavioral meanings to physical world remains poorly understood. Here we provide functional and causal evidence to support a critical role of PPC circuits in this process. We show that PPC neurons exhibit characteristic dynamics related to category learning, adaptively incorporating new sensory information while maintaining stable representations for learned category to choice mappings, a balance in flexibility and stability. Importantly, we provide, to our knowledge, the first evidence supporting that PPC causally contributes to perceptual decision-making in categorizing new sensory information, a key process in category leaning. Furthermore, at the circuit level, we identified a necessary role of PPC to auditory cortex feedback projections, providing a critical foundation for further dissection of circuit-level mechanisms of category learning. After the stimuli were well-learned, PPC continued to represent stimulus categories (Fig. 2) and behavioral choices (6, 11, 13, 23, 24). This is likely to be important for guiding future decisions on new sensory stimuli, and reducing biasing effects from random factors (Fig. 3). Some of recent studies showed that silencing PPC impaired visually guided (14, 17, 27, 28) but not auditory guided (17, 26, 28) choice behaviors in rodents, while in primates, silencing parietal cortex did not influence either visual motion (23) or self motion (24) guided choice behavior, suggesting that that PPC may also play certain visual-specific functions in rodents. Our results, however, support a more general functional role of PPC in perceptual decision-making when facing novel conditions. More recent studies in rodents also using well-trained stimuli showed increased performance in tasks requiring short-term memory or working memory following inactivation of PPC (21, 22), likely attributable to an interference with short-term memory by the effect of PPC on recent choice history, but not an direct effect on perceptual decision-making per se. This is consistent with our observation that PPC activity counterbalances recent history bias. Our results thus provide a framework for mechanistic understanding of neuronal circuit basis of decision-making integrating new sensory information with prior knowledge in a dynamic environment.
Author contributions
L.Z. and N.L.X. conceived the project and designed the experiments. L.Z. performed all the experiments and data analysis. Y.Z. and L.Z. performed the chemogenetic experiments. L.Z. and C.A.D. conceived and performed the GLMM analysis. L.Z., J.P. and N.L.X. developed the hardware and software of the behavior and imaging system. L.Z. and N.L.X. wrote the manuscript with contributions from C.A.D.
References




