

Abstract
Swarmer cells of the gram-negative pathogenic bacteria Proteus mirabilis and Vibrio parahaemolyticus become long (>10-100 μm) and multinucleate during their growth and motility on polymer surfaces. We demonstrate increasing cell length is accompanied by a large increase in flexibility. Using a microfluidic assay to measure single-cell mechanics, we identified large differences in swarmer cell stiffness of (bending rigidity of P. mirabilis, 9.6 x 10 -22 N m2; V. parahaemolyticus, 9.7 x 10-23 N m2) compared to vegetative cells (1.4 x 10-20 N m2 and 3.2 x 10-22 N m2, respectively). The reduction in bending rigidity (∽3-15 fold) was accompanied by a decrease in the average polysaccharide strand length of the peptidoglycan layer of the cell wall from 28-30 to 19-22 disaccharides. Atomic force microscopy revealed a reduction in P. mirabilis peptidoglycan thickness from 1.5 nm (vegetative) to 1.0 nm (swarmer) and electron cryotomography indicated changes in swarmer cell wall morphology. P. mirabilis and V. parahaemolyticus swarmer cells became increasingly sensitive to osmotic pressure and susceptible to cell wall-modifying antibiotics (compared to vegetative cells) — they were ∽30% more likely to die after 3 h of treatment with minimum inhibitory concentrations of the β-lactams cephalexin and penicillin G. Long, flexible swarmer cells enables these pathogenic bacteria to form multicellular structures and promotes community motility. The adaptive cost of swarming is offset by a fitness cost in which cells are more susceptible to physical and chemical changes in their environment, thereby suggesting the development of new chemotherapies for bacteria that leverage swarming for survival.
Significance Statement Proteus mirabilis and Vibrio parahaemolyticus are bacteria that infect humans. To adapt to environmental changes, these bacteria alter their cell morphology and move collectively to access new sources of nutrients in a process referred to as ‘swarming’. We found that a change in the composition and thickness of the peptidoglycan layer of the cell wall makes swarmer cells of P. mirabilis and V. parahaemolyticus more flexible (i.e., reduced cell stiffness) and increases their sensitivity to osmotic pressure and cell-wall targeting antibiotics (e.g., β-lactams). These results highlight the importance of assessing the extracellular environment in determining antibiotic doses and the use of β-lactams antibiotics for treating infections caused by swarmer cells of P. mirabilis and V. parahaemolyticus.
Introduction
Bacteria have evolved a variety of mechanisms to adapt to their physical environment. For example, in response to fluctuating environmental conditions, changes in biochemistry and gene regulation can alter bacterial shape and increase cell fitness. Cell filamentation is a commonly observed change in bacterial cell shape (1, 2) and has been described as a mechanism that enables bacteria to evade predation by the innate immune system during host infections (1).
In close proximity to surfaces, many bacteria alter their morphology and leverage cell-cell physical contact to move collectively to access new sources of nutrients and growth factors (3, 4). Referred to as ‘swarming’, this process is common among motile bacteria, has been connected to bacterial pathogenesis and infections, and is an example of adaptive behavior (3, 4). Swarmer cells of Salmonella enterica, Pseudomonas aeruginosa, Serratia marcescens, and Bacillus subtilis have reduced susceptibility — compared to vegetative cells — to a variety of antibiotics that target protein translation, DNA transcription, and the bacterial cell membrane and cell wall (5-8). The specific biochemical and biophysical mechanisms underlying these observations are unknown.
Here, we describe physical changes in swarmer cells of the gram-negative pathogenic bacteria Proteus mirabilis and Vibrio parahaemolyticus that increase their susceptibility to cell wall-targeting clinical antibiotics. We found that large changes in the length of P. mirabilis and V. parahaemolyticus swarmer cells are accompanied by an increase in flexibility (i.e., a reduction in cell stiffness) that enables long cells to pack together tightly and form cell-cell interactions; these interactions have been demonstrated previously to promote surface motility (9). Using biophysical, biochemical, and structural techniques, we quantified changes in the structure and composition of the P. mirabilis and V. parahaemolyticus cell wall in swarmer and vegetative cells and characterized their susceptibility to osmotic changes and cell wall-modifying antibiotics. Our results indicate that morphological changes that enable these bacteria to adapt to new physical environments come at a significant fitness cost: cells become more susceptible to changes in their chemical environment. These results predict cell wall-modifying antibiotics should deter infections of P. mirabilis and V. parahaemolyticus in which swarming is implicated (e.g., in urinary tract infections).
Results
The bending rigidity of P. mirabilis and V. parahaemolyticus cells decreases during swarming
During surface motility, P. mirabilis and V. parahaemolyticus cells grow into swarmers that are characteristically long (10-100 μm) and present a high surface density of flagella that enables them to translate through viscous environments (3, 10). We found that these swarmer cells display an unusual phenotype that is rarely observed among gram-negative bacteria: cells become remarkably flexible and their shape is dynamically altered by adjacent cell motion and collisions (Fig. 1). The ability of P. mirabilis swarmer cells to form cell-cell contacts plays a role in their cooperative motility (10), leading us to hypothesize that increasing flexibility enables these long cells to optimize packing into multicellular structures that move cooperatively across surfaces. Bacterial cell mechanics is generally attributed to the peptidoglycan (PG) layer of the cell wall, which has a thickness of ∽3-50 nm and surrounds the cytoplasmic membrane (11). Very little is known about mechanical regulation in bacteria (12-17) and we are unaware of studies connecting swarming to changes in cell mechanics. We quantified changes in swarmer-cell stiffness using cell-bending assays in a reloadable, poly(dimethylsiloxane) microfluidic system (Fig. 2 and S1) that is related to a method developed previously (18). In bending assays, we applied a shear fluid force to multiple filamentous cells, resulting in horizontal deflection of their cell tips (Fig. 2); fitting the deflection data to a mechanical model provided us with a value of the (flexural) bending rigidity of cells (Fig. S2). Introducing a reloadable mechanism enabled us to perform rapid bending measurements of P. mirabilis and V. parahaemolyticus swarmer cells after isolating them from swarm plates. As a point of comparison, we filamented vegetative cells of P. mirabilis and V. parahaemolyticus using aztreonam — an inhibitor of the division specific transpeptidase PBP3 — to match the length of swarmer cells and compared their bending rigidity values to swarmer cells. As a control, we measured the bending rigidity of cells of Escherichia coli strain MG1655 that we filamented using aztreonam, and determined the value to be 9.7 x 10-23 N m2 (Fig. 3); using a value for the thickness of the PG of 4-nm (19) yields a Young’s modulus of 26 MPa, which is close to values that have been reported previously (12, 18).

Images demonstrating the flexibility of P. mirabilis and V. parahaemolyticus swarmer cells. (A) Time-series of P. mirabilis swarmer cells in a colony actively moving across the surface of a 1.5% agarose gel. A representative cell, false colored green, has a generally straight shape at t = 0 s and is bent in half at t = 0.98 s. Most of the cells in this frame are bending during this imaging sequence. (B) A time-series of V. parahaemolyticus swarmer cells in a colony actively moving across the surface of a 1.4% agarose gel. A representative cell (false colored purple) has a generally straight shape at t = 0 s and is bent in half at t = 4.02 s.

Using a reloadable microfluidic-based assay to determine bacterial cell stiffness. (A) Schematic of the microfluidic channel used to apply a user-defined shear force to bend filamentous or swarmer cells. Single-sided green arrows depict the flow of fluid through the central channel; the parabolic flow profile of the fluid is shown. Double-sided green arrows indicate the vacuum chamber used to load cells into side channels and to empty the device. (B) Cartoon of a flexible bacterium (left) and a stiff bacterium (right) under flow force (Vflow). xmax indicates the deflection of cells in the flow; 2r = cell diameter; L = cell length in contact with the flow force. (C) Representative images of filamentous cells of P. mirabilis in no flow (left) and flow (right) conditions (top) and P. mirabilis swarmers (bottom). Purple dashed lines indicate the position of a cell tip under no flow conditions and black dashed lines illustrate the position after flow is applied using a gravity-fed mechanism. The arrow indicates the direction of fluid flow in the channel.

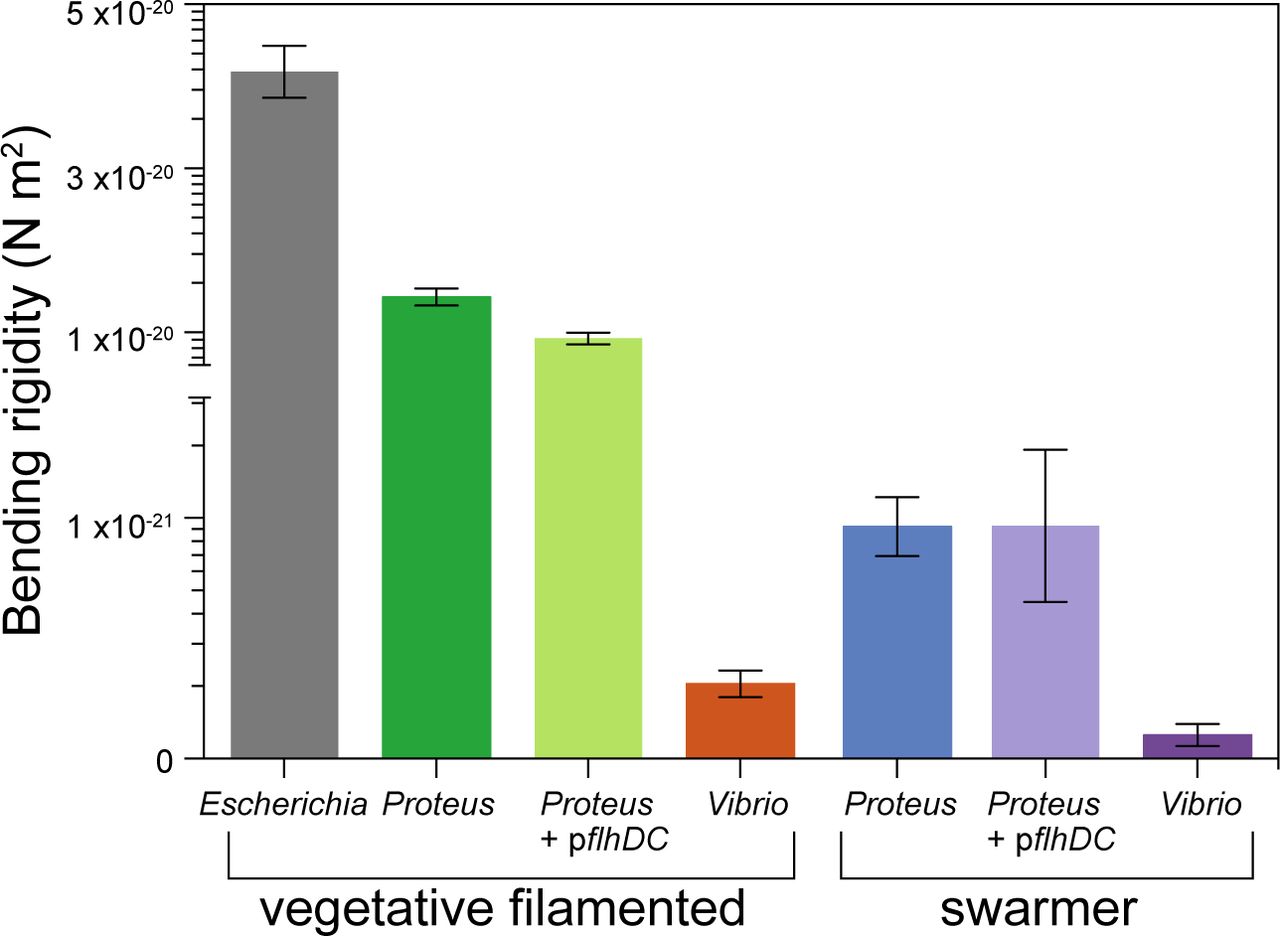
P. mirabilis and V. parahaemolyticus swarmer cells have a lower bending rigidity than vegetative cells. We measured the bending rigidity of P. mirabilis and V. parahaemolyticus swarmer cells and filamentous vegetative cells in a microfluidic flow assay and included measurements of vegetative E. coli cells. P. mirabilis swarmers exhibited 15-fold lower bending rigidity than vegetative cells; V. parahaemolyticus swarmers were 3-fold less rigid than vegetative cells. Overexpression of FlhDC (from the plasmid-encoded pflhDC) had little effect on the stiffness of P. mirabilis vegetative and swarmer cells. Error bars represent the 95% confidence interval of a fit to data. n > 100 cells, from at least 3 independent experiments.
We used bending rigidity as a parameter to quantify P. mirabilis and V. parahaemolyticus cell stiffness rather than Young’s modulus, as the latter metric of the mechanical properties of materials is dependent on cell-wall thickness, which we hypothesized may change during swarming (described below). We found a substantial decrease in the bending rigidity of swarmer cells of both P. mirabilis (∽15-fold) and V. parahaemolyticus (3-fold) compared to their vegetative cell counterparts (Fig. 3), which is consistent with our observations of the shape and behavior of these cells during swarming on surfaces. V. parahaemolyticus vegetative cells were remarkably flexible: ∽134-fold more so than E. coli cells and ∽3-fold more than P. mirabilis swarmer cells (Fig. 3).
To confirm that using aztreonam to inhibit PBP3 and produce filamentous cells did not change the cross-linking density of PG at the division plane and alter cell mechanics, we compared bending rigidity values of cells treated with aztreonam and cells filamented by overexpressing SulA, a protein that prevents polymerization of the division protein FtsZ and blocks cell division. Both mechanisms of filamenting E. coli cells produced similar bending rigidity values: 4.0 x 10-20 N m2 (SulA) and 4.1 x 10-20 N m2 (aztreonam) (Fig. S4A). To account for the high surface density of flagella on swarmer cells, which may contribute to cell drag in fluid flow and bias measurements, we performed bending rigidity measurements (Fig. S4B) on two K – 12-derived strains of E. coli with substantially different flagella densities (Fig. S4C) and observed no appreciable change in values (bending rigidity of 4.0 x 10-20 N m2 at high flagella density and 4.1 x 10-23 N m2 at low flagella density; Fig. S4B).
Overexpressing FlhDC — the heterohexameric activator that is important for swarming — in vegetative cells growing in liquid produces a phenotype that replicates many of the characteristics of swarmer cells, including increased cell length and flagella density (20). The relationship between FlhDC and cell mechanics has not yet been reported. To test whether FlhDC is connected to changes in swarmer cell stiffness, we overexpressed the protein from the plasmid-encoded genes pflhDC in filamentous cells of P. mirabilis and measured their bending rigidity. We detected a ∽1.5-fold difference in bending rigidity between wildtype (1.4 x 10-20 N m2) and pflhDC-containing P. mirabilis vegetative cells (9.2 x 10-21 N m2), and no difference compared to pflhDC-containing P. mirabilis swarmer cells (Fig. 3). These results indicate that FlhDC overexpression during swarming contributes a small amount to the mechanical phenotype of swarming cells, however the majority of the mechanical changes we observe arises from PG alterations through another regulatory pathway.
Changes in PG composition of P. mirabilis and V. parahaemolyticus swarmer cells
PG consists of the disaccharide building block β-(1,4)-N-acetylmuramic acid / N-acetyl-glucosamine (MurNAc-GlcNAc) in which a pentapeptide is attached to each 3’-OH group on MurNAc. Cross-linking between adjacent pentapeptides creates a mesh-like polymeric layer, and altering its structure and composition affects cell-mechanical properties (14, 15). To determine whether the PG composition of P. mirabilis and V. parahaemolyticus cells changes during swarming, we isolated PG sacculi from vegetative and swarmer cells and used ultra performance liquid chromatography-mass spectrometry (UPLC-MS) to quantify its chemical composition (Fig. S5). As the PG composition of V. parahaemolyticus has not yet been reported, we characterized its muropeptide stem peptide using UPLC – MS / MS (Fig. S6 and Table S1). Similar to E. coli (21) and P. mirabilis (22),V. parahaemolyticus has a PG structure that is conserved across other gram-negative bacteria, in which the peptide stem consists of L-Ala-D-Glu-meso-diaminopimelic acid (meso-DAP)-D-Ala-D-Ala (Fig. S6).
Compared to vegetative cells, P. mirabilis swarmer cells contained fewer monomers (MurNAc-GlcNAc), more dimers, and more anhydrous-containing saccharides, which are found at the terminating end of glycan polymers (Fig. 4A) (11). We detected no differences in the relative abundance of trimers between swarmer and vegetative cells of P. mirabilis (Fig. 4A). The increase in anhydrous-containing saccharides that we observed in P. mirabilis swarmer cells was correlated with a decrease in polysaccharide length (Fig. 4B; the values plotted reflect the number of MurNAc-GlcNAc dimers). A similar increase in anhydrous-containing saccharides and decreased length of polysaccharides occurred in V. parahaemolyticus swarmer cells (Fig. 4A, B). We found no change in cross-linking density between vegetative and swarmer cells of either P. mirabilis or V. parahaemolyticus (Fig. 4C).
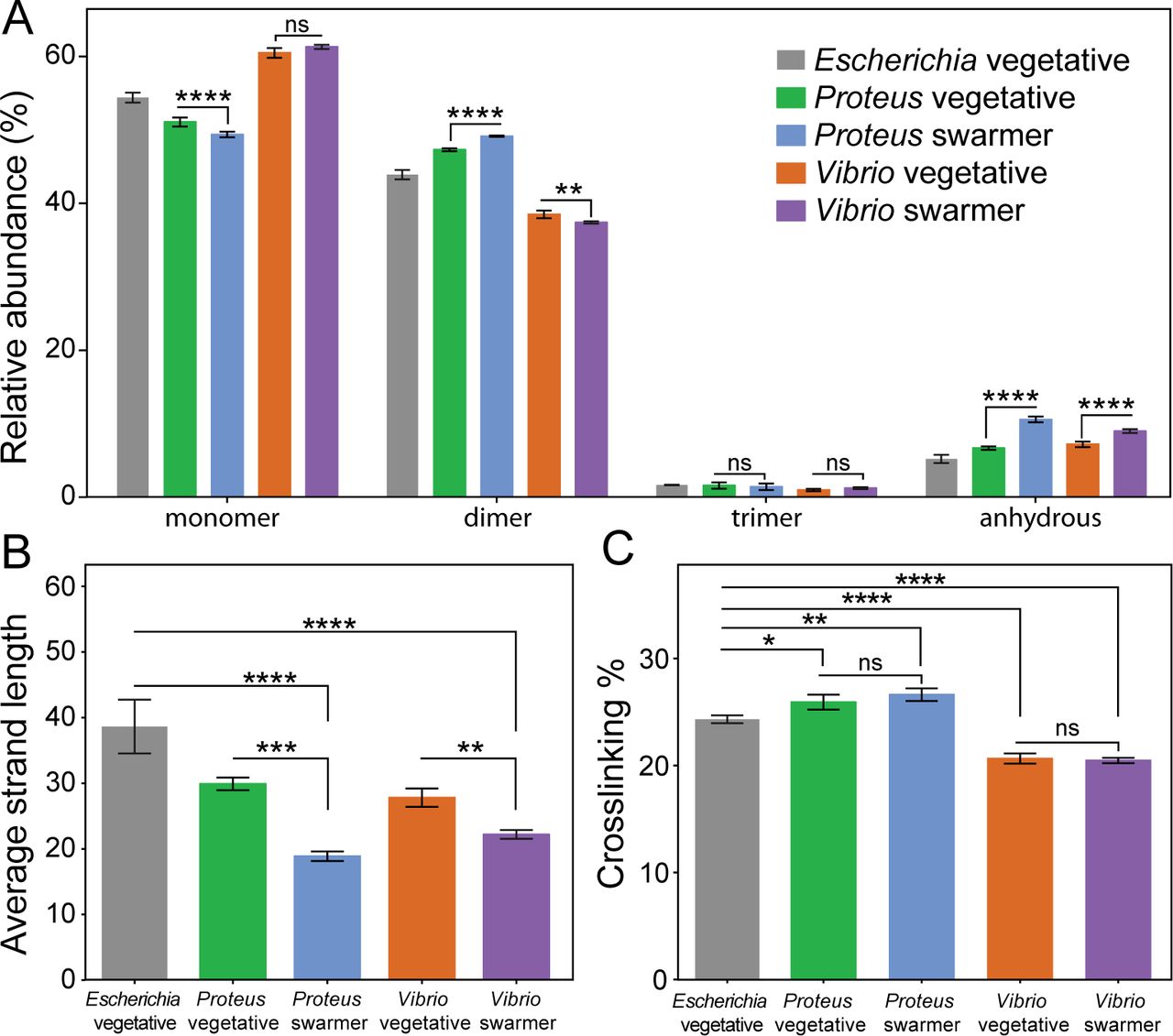
Alterations in the PG muropeptide composition of P. mirabilis and V. parahaemolyticus swarmer cells. (A) UPLC-MS data reveal that the muropeptide composition of P. mirabilis and V. parahaemolyticus vegetative and swarmer cells differs slightly in the abundance of monomer, dimer, and anhydrous-terminated saccharides. (B) We observed a relative increase in the amount of anhydrous-containing saccharides in swarmers consistent with a decrease in polysaccharide strand length. (C) There was no change in PG cross-linking of P. mirabilis and V. parahaemolyticus vegetative and swarmer cells, although V. parahaemolyticus does display a lower level of cross-linking. n = 3 biological replicates. Error bars represent the standard deviation of the mean. For (A-C), significance was determined via two-way analysis of variance: *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001, ns = not significant (P > 0.05).
Swarmer cells have a reduced PG thickness and display membrane defects
Changes in the thickness of the PG layer and structure of the cell envelope may also explain the observed decrease in swarmer cell stiffness. To identify changes in PG thickness of swarmer cells, we isolated the intact layer of PG (i.e., sacculi) from P. mirabilis vegetative and swarmer cells and measured the thickness of dried sacculi using tapping-mode atomic force microscopy (AFM) (Fig. 5A). Differences in the nanoscopic appearance of the sacculi of different cells were not observed by AFM (Fig. S7). The thickness of isolated, dry P. mirabilis swarmer cell sacculi (1.0 ± 0.2 nm) was reduced ∽1.5-fold compared to vegetative cells (1.5 ± 0.2 nm) (Fig. 5A). V. parahaemolyticus swarmer cells (0.6 ± 0.1 nm) exhibited a similar ∽1.2-fold decrease in thickness compared to vegetative cells (0.8 ± 0.2 nm). Earlier AFM measurements of isolated sacculi indicated that dehydration reduced the thickness of Escherichia coli PG by ∽2x, which we used to estimate the dimensions of hydrated PG from P. mirabilis (3.1 and 2.1 nm for vegetative and swarmer cells, respectively) and V. parahaemolyticus (1.7 and 1.4 nm, respectively) (23). A comparison of PG thickness and cell bending rigidity suggested that the relationship between these data is approximately exponential (R2 = 0.9874) (Fig. 5B).

AFM reveals the PG layer of P. mirabilis and V. parahaemolyticus swarmer cells is thinner than in vegetative cells and ECT demonstrates a reduced membrane-to-membrane distance. (A) Sacculi were isolated from cells, dried, and imaged by AFM. The thickness of swarmer cell PG was reduced compared to vegetative cells. We analyzed >65 vegetative cells of E. coli, P. mirabilis, and V. parahaemolyticus, >65 P. mirabilis swarmer cells, and 7 V. parahaemolyticus swarmer cells. Error bars represent the standard deviation of the mean. (B) Bending rigidity and cell wall thickness display an approximately exponential relationship (R2 = 0.9874). (C) Sub-tomogram-averaged ECT volume images of the P. mirabilis vegetative (left) and swarmer (right) cell wall. Two central slices of sub-tomogram average volume images with normalized image densities are shown. Yellow dashed line indicates the orientation used for gray-value measurements. ES, extracellular space; OM, outer membrane; PG, peptidoglycan; IM, inner membrane; CP, cytoplasm. (D) The density profile of sub-tomogram-averaged ECT volume images reveals a reduced membrane-to-membrane distance in swarmer cells. The vertical axis is the normalized gray value with the darkest value equal to 0 and the lightest value equal to 1. The red dashed line denotes the average gray value of the extracellular space and serves as a reference for the background; the blue shaded area indicates the thickness of the putative PG layer.
A caveat with conversions between dried and hydrated values is that they will be most accurate for PG that best mimics the structure and composition (e.g., crosslinking and polysaccharide composition) of the reference material: E. coli PG. Alterations in the polysaccharide length of PG from P. mirabilis and V. parahaemolyticus may be more elastic and stretch out during drying, thereby appearing to have a thickness that is reduced compared to E. coli PG. Our control measurements on isolated, dry E. coli sacculi yielded a thickness of 2.0 nm, which varies from the value of 3.0 nm published by Yao et al. using the same technique (23). A difference of ∽30% between these E. coli measurements may arise for several reasons, including: variations in physical conditions that can impact AFM measurements, improvement in the resolution of AFMs, and / or the precision of fitting force curves.
The relatively low variability in the values we measured for isolated sacculi from P. mirabilis and V. parahaemolyticus cells (∽16-25%) demonstrated a consistent reduction in PG thickness for swarmer cells, suggesting that our measurements are sufficient for comparing PG from vegetative and swarming cells, and demonstrating a connection between PG and changes in cell mechanics. In contrast, the variability in AFM measurements and analysis makes us less comfortable comparing absolute values of PG thickness to those reported for bacteria in other papers.
To complement AFM measurements, we sought to measure the thickness of native PG using electron cryotomography (ECT) on intact vegetative and swarmer cells (Figs. 5C,D, S8, and S9). Although we were unable to resolve the thickness of native PG, sub-tomogram-averaged ECT volumes of the P. mirabilis cell wall (Fig. 5C) indicated that the distance between the inner and outer membrane of swarmer cells was smaller than in vegetative cells (Figs. 5C,D). P. mirabilis vegetative cells had a characteristically smooth membrane (Fig. S8A,B) that was similar to the presentation of the membrane found along the lateral, cylindrical walls of swarmer cells (Fig. S8C,D). In contrast, the polar regions of both P. mirabilis and V. parahaemolyticus swarmer cells had an undulating outer membrane suggestive of an altered structure (Fig. S8E,F and S9D-F), and V. parahaemolyticus cells had significant defects in their cell-envelope, including, membrane budding, vesicle formation, and ruptured cell walls (Fig. S9D-F).
P. mirabilis and V. parahaemolyticus swarmer cells are more sensitive to osmotic stress than vegetative cells
To complement cell stiffness measurements, we investigated the mechanics of swarmer cells and filamented vegetative cells by measuring their changes in length and volume in response to changes in osmotic pressure, which will cause cells to shrink cells (in H2O) and swell (in a NaCl solution) (Fig. S10). We filamented P. mirabilis, V. parahaemolyticus, and control E. coli cells using aztreonam to create a range of cell lengths that matched the lengths of P. mirabilis and V. parahaemolyticus swarmer cells. We expected that low values of bending rigidity and compositional and structural changes in PG would cause swarmer cells to elongate in response to changes in osmotic pressure (in comparison to filamented, vegetative cells). We found that P. mirabilis and V. parahaemolyticus swarmer cell extension in response to osmotic shock was significantly larger than filamented, vegetative cells (Fig. 6). To circumvent the cellular production of osmolytes to protect cells from large changes in osmotic pressure during shock (typically produced within 1 min) (24), we used a microfluidic device to rapidly switch between H2O and a NaCl solution and measured changes in cell length before cells adapted (Fig. S10).
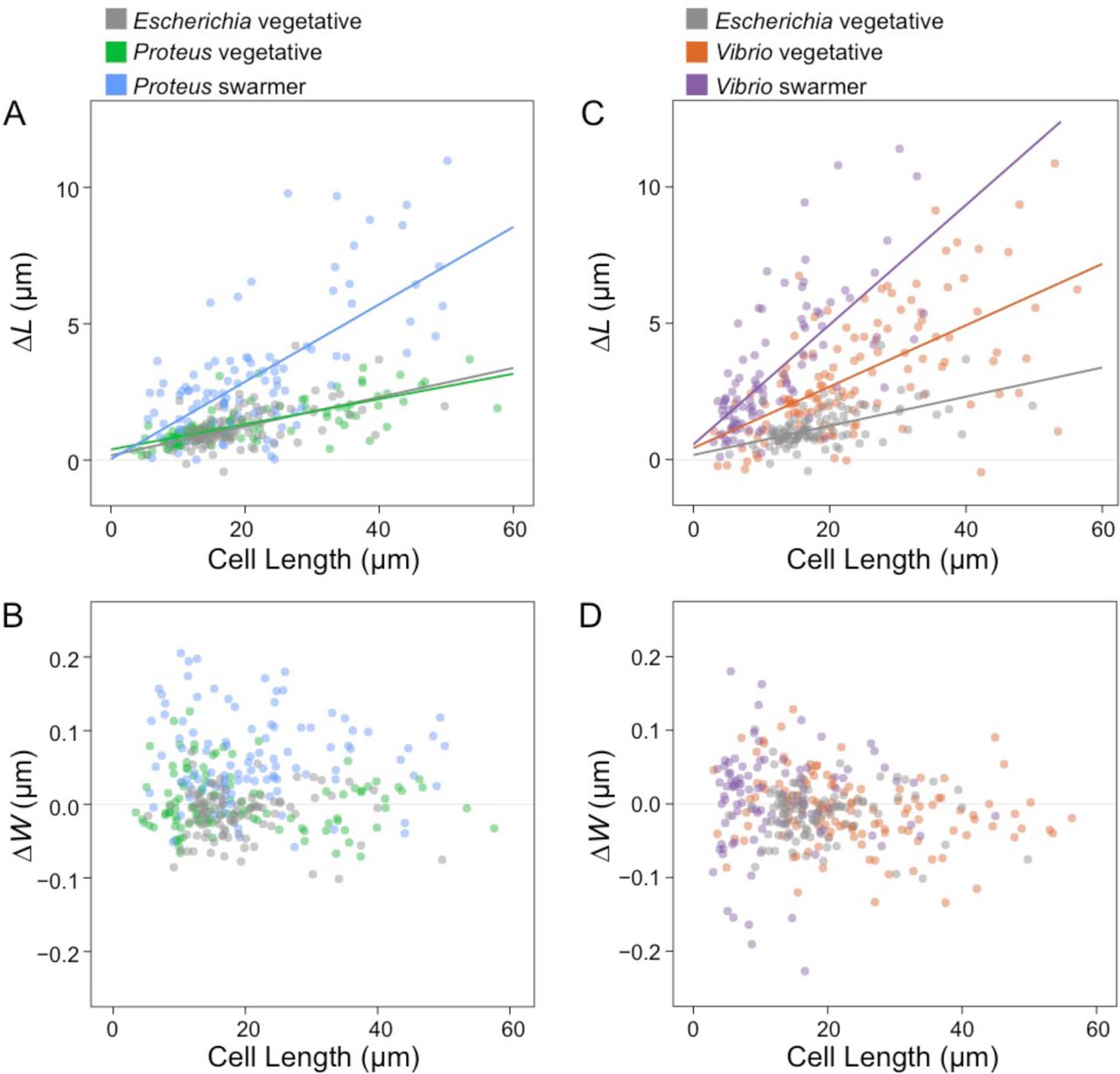
Swarmer cells increase in cell extension during osmotic shock. We calculated ΔL as (cell length in water – cell length in 1 M NaCl) and performed a similar calculation for ΔW, substituting cell width. Cell length indicates length prior to osmotic shock. We filamented all vegetative cells using aztreonam to grow them to lengths that were comparable with P. mirabilis and V. parahaemolyticus swarmers. Lines indicate linear fits to single-cell measurements (circles) of n > 100 cells from at least 3 independent experiments. (A) P. mirabilis swarmer cells have an increase in extension (ΔL) under osmotic shock compared to E. coli and P. mirabilis vegetative cells. (B) V. parahaemolyticus vegetative and swarmer cells have an increase in extension (ΔL) under osmotic shock compared to E. coli. (C) P. mirabilis swarmer cells have an increase in ΔW compared to E. coli; P. mirabilis vegetative cells display a slight decrease in width and increased cell length. (D) There was no observable change in ΔW of V. parahaemolyticus swarmer and vegetative cells.
Osmotic shifts produced small changes in the length (Fig. 6A) and width (Fig. 6B) of vegetative filamented P. mirabilis cells compared to E. coli cells (Figs. 6A, B). In contrast, V. parahaemolyticus vegetative cells substantially increased in cell length (Fig. 6C) compared to E. coli cells, with no observable change in cell width (Fig. 6D). In response to osmotic upshifts (i.e., transitioning from H2O to NaCl), P. mirabilis and V. parahaemolyticus swarmer cells dramatically increased in cell length (Figs. 6A, C) (and to a lesser extent, cell width for P. mirabilis but not for V. parahaemolyticus; Figs. 6B, D) compared to vegetative cells. These results suggest that changes in P. mirabilis and V. parahaemolyticus swarmer cell mechanics make them more responsive to osmotic changes.
Increased susceptibility of P. mirabilis and V. parahaemolyticus swarmers to the cell wall-targeting antibiotics
Although swarming colonies of bacteria display resistance to many antibiotics (5), our experiments suggest P. mirabilis and V. parahaemolyticus swarmers may have increased susceptibility to cell wall-targeting antibiotics that further alter PG structure, composition, and properties. To determine the sensitivity of P. mirabilis and V. parahaemolyticus swarmers to the cell wall-targeting antibiotics cephalexin (inhibits PBP3 and 4) (25) and penicillin G (inhibits PBP3, 4, and 6), we measured swarmer single-cell growth in the presence of the minimum inhibitory concentration (MIC) of antibiotics in a microfluidic device (Figs. S11) for 3 h. The time scale of this experiment was chosen based on two previous reports: 1) treating cells with cephalexin for 3 h was sufficient to kill ∽100% of a population of E. coli cells (26); and 2) ∽2 h of cephalexin treatment was sufficient to observe cell lysis of E. coli cells measured by single cell growth (27).
Using a microfluidic device enabled us to supply cells with a source of continuously replenished fresh growth media to ensure exponential cell growth and a constant concentration of antibiotics. We found that the survival of P. mirabilis (66%) and V. parahaemolyticus (64%) vegetative cells treated with 1X MIC of cephalexin was slightly higher than E. coli cells (55%) (Figs. 7A, S12). Treating P. mirabilis and V. parahaemolyticus swarmer cells with 1X MIC of cephalexin reduced survival to 37% and 19%, respectively (Figs. 7A, S12), indicating their increased susceptibility. Rates of cell survival in the presence of penicillin G were similar to the use of cephelexin (Fig. 7A). We characterized dead cells using microscopy to measure membrane blebbing, cell lysis, and changes in the refractive index of cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Swarmer cells are more susceptible to antibiotics that target the cell wall than are vegetative cells. (A) Survival of cells treated with 1X MIC of cephalexin after 3 h of incubation. We define percent survival as (cell countno lysis / cell counttotal) x 100. P. mirabilis and V. parahaemolyticus swarmer cells exhibit decreased levels (∽30%) of survival compared to vegetative cells; n ≥ 90 cells from at least two independent experiments. A similar decrease occurred when P. mirabilis was treated with penicillin G; n ≥ 77 cells from at least two independent experiments). Error bars represent the standard deviation of the mean. (B) After exposure to cephalexin or penicillin G, the survival time of P. mirabilis and V. parahaemolyticus swarmers was 2∽3 fold lower than for vegetative cells. Survival time was determined for ≥ 49 cells that lysed from at least 2 independent experiments. Significance in A-B was determined two-tailed t-test: *P ≤ 0.05, **P ≤ 0.01, ****P ≤ 0.0001.
Cell wall-targeting antibiotics are most effective against actively growing cells (28, 29) and therefore increases in growth rate will reduce cell survival. To determine whether a growth rate phenotype explains our observations, we normalized data for cell length, and were unable to detect a change in the growth rate of swarmer cells when they were treated with these two antibiotics (Fig. S13). We measured the mean survival time of cells (the amount of time elapsed after treatment with drugs before cell death) for P. mirabilis and V. parahaemolyticus swarmer cells treated with cephalexin and penicillin G (Fig. 7B) and found that the survival time for these swarmer cells was lower than that for vegetative cells (Fig. 7B), which is consistent with alterations in the PG.
Discussion
P. mirabilis is commonly associated with complicated urinary tract infections and increased mortality in cases of bacteremia (30). Swarming is hypothesized to enable P. mirabilis cell movement from the ureter to the kidney; in support of this hypothesis, swarming deficient mutants have lower rates of host infection (31). We found that during swarming, P. mirabilis and V. parahaemolyticus cells become more flexible due to changes in PG composition and cell wall structure. Presumably this phenotype conveys an adaptive advantage for cells that we hypothesize improves community motility by enabling long swarmer cells to form cell-cell contacts that are demonstrated to enhance motility. The adaptive advantage of swarming in P. mirabilis and V. parahaemolyticus is offset by a decrease in their fitness, as cells become more sensitive to osmotic changes and cell wall-targeting antibiotics, thereby creating an “Achilles heal” for targeting this phenotype in infectious diseases. This study highlights the plasticity of bacteria (1), the need for devising tests for treatments that measure efficacy against cells in a physiological state relevant to specific infections, and re-considering the use of cell wall-targeting antibiotics for treating UTIs in which P. mirabilis may be present.
Materials and Methods
Bacterial strains and cell culture
P. mirabilis strain HI4320, V. parahaemolyticus LM5674, Escherichia coli MG1655 (CGSC #6300), and plasmids pflhDC (10) and psulA (18) were used for experiments used in this paper. P. mirabilis was grown in PLB nutrient medium consisting of 1% peptone (weight / volume), 0.5% yeast extract (w / v), and 1% NaCl (w / v). V. parahaemolyticus was grown in nutrient medium (HI medium) consisting of 2.5% heart infusion (w / v) and 2.5% NaCl (w / v). E. coli was grown in lysogeny broth (LB) consisting of 1% tryptone (w / v), 0.5% yeast extract (w / v), and 1% NaCl (w / v). All strains were grown at 30 °C with shaking at 200 rpm. Additional data and methods are described in Supplementary Information.
Acknowledgements
We thank Linda McCarter for V. parahaemolyticus LM5674, Suckjoon Jun for the psulA plasmid, Cameron Scarlett and Molly Pellitteri-Hahn for mass spectrometry support, and Julie Last for technical assistance with AFM measurements. This research was supported by NIH grant 1DP2OD008735-01, National Science Foundation grant DMR-1121288, a Mao Wisconsin Distinguished Graduate Fellowship (to M.R.), and an NSF postdoctoral fellowship (#1202622 to P.M.O), and the Howard Hughes Medical Institute.
References




