

Abstract
Human capacity to remember experienced episodes over a long period of time has its roots in childhood and develops throughout the lifespan. However, the neural regions supporting memory consolidation in the developing brain remain to be ascertained. The present study examined system-level memory consolidation of object-location associations after one night of sleep (short delay) and after two weeks (long delay), and its relation to structural brain measures in normally developing term born and preterm born 6-year-old children, as well as in young adults as a reference group of mature consolidation systems. We showed that final learning performance was reduced in preterm in comparison to term born children, who in turn were outperformed by young adults. There were no differences in short- and long-delay memory consolidation between term and preterm born children. Despite comparable short-delay memory consolidation in all groups, both term and preterm born children showed less efficient long-delay memory consolidation in comparison to young adults. Moreover, long-delay memory consolidation was positively associated with larger hippocampal volume in children, while a thinner medial orbitofrontal cortex was associated with better overall memory retention rates in all age groups. Thinner medial orbitofrontal cortex was furthermore associated with higher final learning performance in children. Taken together, the results suggest that temporal dynamics of memory consolidation and its association with structural brain measures in 6-year-old term born and preterm born children are comparable but differ from young adults.
RESEARCH HIGHLIGHTS
Short-delay memory consolidation of object-location associations does not differ between 6-year-old preterm born and term born children, as well as between children and young adults.
Long-delay memory consolidation in 6-year-old children is less efficient than in young adults.
Larger HC volume is associated with higher long-delay retention rates in 6-year-old children, while thinner medial OFC predicted higher memory retention rates in all groups.
Thinner medial OFC is associated with higher final learning performance in 6-year-old children and adults.
1 INTRODUCTION
Humans have an impressive capacity to store and retrieve memories of past experiences with unique temporal-spatial features for years and even decades (Squire et al., 2015; Tulving & Markowitsch, 1998; Tulving, 2002). This is possible due to memory consolidation, a dynamic and complex process through which acquired memory traces become long-lasting (Dudai, 2004; Dudai et al., 2015; Moscovitch & Gilboa, 2021). The interplay between the hippocampus (HC) and neocortical brain structures underlies the formation, maintenance, and retrieval of declarative memory (Moscovitch & Gilboa, 2021; Simons & Spiers, 2003; Squire et al., 2015b; Squire & Alvarez, 1995). Events are bound by the HC into a coherent memory trace containing contextually rich information that is distributed within neocortical areas (Eichenbaum, 2017; Nadel & Peterson, 2013). The ventromedial prefrontal cortex the (vmPFC) is involved in the evolvement of memory representations either by integrating new memory representations into pre-existing schemata or by forming new schemata (Frankland & Bontempi, 2005; Squire et al., 2015b; Tompary & Davachi, 2017; Tse et al., 2007, 2011). Taken together, memory consolidation involves an intricate, temporally evolving interplay of different brain regions, networks, and cognitive processes to enable long-term retrieval of past experiences.
1.1 Memory consolidation across development
From an ontogenetic perspective, the ability to retrieve long-term declarative memories emerges with the close of childhood amnesia, i.e., the inability to recollect early life events, around four to seven years of age (Alberini & Travaglia, 2017; Hayne, 2004). This indicates the kickoff of long-term memory stabilization (Bauer, 2006; Henke, 2010; Morgan & Hayne, 2011; Perner & Ruffman, 1995; Scarf et al., 2013; Tustin & Hayne, 2010). In Germany the transition from kindergarten to school occurs at the age of 6 years and is accompanied by dramatic increase in learning and knowledge accumulation, demands posed on children due to a structured primary school curriculum (Linberg et al., 2013; Mashburn et al., 2018). It challenges not only cognitive abilities but potentially also memory consolidation processes in this age group (Brod et al., 2017; Nolden et al., 2021). Although successful retrieval of complex memory representations starts to steadily improve at the age of 5 to 6 years (Drummey & Newcombe, 2002; Riggins, 2014; Sluzenski et al., 2006), the rate of memory retention across longer consolidation periods (Murre & Dros, 2015; Ebbinghaus, 1885; Müller & Pilzecker, 1900) is much less researched and may progress with different temporal dynamics in children in comparison to adults. For instance, it has been shown that short-delay memory consolidation rates (i.e., measured after one night of sleep) were comparable between children aged 6 to 8 years and young adults for word-pair associates (Wilhelm et al., 2008), and were even more efficient in 7 to 12 years old children in comparison to young adults for non-objects and their functions (Peiffer et al., 2020a) and for “what-where-when” memories of faces (Wang et al., 2018). At the same time, findings show that in children, successful retrieval of events over a long time increases with increasing age (Boyer et al., 1994; Østby et al., 2012a; Peterson, 2002). However, little is known about memory consolidation of complex representations over shorter and longer delays in 6-year-old children who are about to start or are just starting school, as it has not yet been systematically investigated in the field of developmental neuroscience.
The core neural structures involved in memory consolidation undergo profound morphological changes in children. For instance, the HC reaches its relative maturity in middle childhood (Keresztes et al., 2017; Shing et al., 2010; Uematsu et al., 2012, but also see Lee et al. 2016), which is concomitant with an improvement in the ability to bind event features into a coherent representation (Lee et al., 2016; Shing et al., 2008, 2010; Sluzenski et al., 2006). On the other hand, memory control processes (e.g., strategy use, resolving mnemonic interferences) that benefit encoding, consolidation, and later retrieval of memories depends more on the ventrolateral prefrontal cortex (vlPFC; Badre & Wagner, 2007; Kuhl et al., 2012), while schemaintegration processes that benefit long-term consolidation depend more on the vmPFC (Brod & Shing, 2018). Each of these regions shows more prolonged maturation and reaches its relative maturity only in late adolescence/young adulthood (Gogtay et al., 2004a; Muftuler et al., 2012; Shing et al., 2010a; Sousa et al., 2018; Uda et al., 2015; Waber et al., 2007). In an exceptional study that examined consolidation and brain correlates in participants aged 8 to 19 years, Østby et al. (2012a) showed that a thinner orbitofrontal cortex (OFC) was associated with a higher shortdelay memory retention, while bigger HC volumes were related to higher memory retention rates after 1-week delay in a visuospatial task. Taken together, these findings indicate that extended developmental trajectories of HC and PFC maturation may affect the efficiency of memory consolidation processes in children, beyond their effects on encoding or retrieval (Shing et al., 2008b, 2010b).
1.2 Preterm birth and neurocognitive development
Prenatal and perinatal conditions can alter the developing brain (Buss et al., 2012), among which premature birth has been shown to greatly affect brain and neurocognitive development (Nosarti & Froudist-Walsh, 2016). Approximately 11% of children are born prematurely (i.e., born before the completion of 37 weeks of gestation; WHO, 2021) annually worldwide (Walani, 2020). Premature birth is associated with altered structural brain anatomy in regions critical to memory consolidation – i.e., reduced grey matter (GM) and white matter (WM) volumes in temporal and frontal brain regions (Ball et al., 2012; Nosarti et al., 2014a; Nosarti & Froudist-Walsh, 2016). Reduced HC volume in preterm born individuals has been observed from infancy through childhood, adolescence, and into adulthood (Aanes et al., 2020; Arhan et al., 2017; Ball et al., 2012; de Kieviet et al., 2012). A growing body of research suggests that brain injury or altered neurodevelopmental trajectories due to premature birth are related to impaired cognitive functioning (e.g., information processing speed, verbal abilities; Linsell et al., 2015; Murray et al., 2014; Rose et al., 2011; Stålnacke et al., 2019), executive functioning (e.g., working memory, inhibition; Arpi et al., 2019; Beauchamp et al., 2008; Ford et al., 2011), and spatial learning (Cimadevilla et al., 2014). Such deviation in brain functioning may pose a special challenge for preterm born children during the transition from kindergarten to school.
Evidence for the effect of preterm birth on memory functioning in children is more mixed. For example, preterm born preschool children showed impaired verbal short-term memory in comparison to their term born peers (Baron et al., 2012; Dall’Oglio et al., 2010; Roberts et al., 2011). It was also reported that 7-year-old preterm born children showed imparied learning and immediate spatial and visual memory (Thompson et al., 2013a, 2014) and deficits in learning and immediate and delayed (20 minutes) visuo-spatial memory (Omizzolo et al., 2014). In addition, Quesada et al. (2014) reported poorer general and visual memory in preterm born children aged 6 to 10 years. On the other hand, no deficits in delayed (20 minutes) visuo-spatial recognition (Omizzolo et al., 2014a) for 7-year-old preterm born children was reported as well as no impairment in delayed verbal and visual recognition in preterm born children aged 6 to 10 years (Quesada et al., 2014). Taken together, current empirical evidence suggests inconsistent patterns of memory functioning in children born preterm. Of note is that preterm born cohorts examined in previous research are often very heterogenous, including individuals with various perinatal complications (e.g., insults, intraventricular hemmorage, etc.) and known learning disabilities. The majority of studies did not apply inclusion criteria for general cognitive functioning (e.g., IQ), which may confound the results comparing preterm born cohorts with typically developing term born children (Fernández de Gamarra-Oca et al., 2021). Importantly, temporal dynamics of long-term memory consolidation in 6-year old preterm born children has not yet been systematically characterized.
Therefore, thus far it is unclear how efficient long-term memory consolidation is in term born and preterm born children from early and middle childhood, a developmental stage during which brain regions underlying the ability to bind events into a coherent representation, to store memories, and to use strategies and exert control over memory retrieval, undergo profound changes (Lenroot & Giedd, 2006; Mills et al., 2016; Ofen et al., 2007; Shing et al., 2010a). In this study, we examined the consolidation of object-location associations across a 2-week-period, comparing 6-year-old full term born and preterm born children to young adults serving as a reference group with a mature memory consolidation system. To counteract the sample heterogenity that may confound previous research on the long-term neurocognitive consequences of premature birth and increase sample comparability, we included only typically developing term born and preterm born children without further birth complications and with at least average IQ. Using an object-location association task, we tracked initial learning of new information up to a predetermined criterion, as well as memory consolidation thereafter following a short delay (after one night of sleep) and a long delay (after two weeks). To minimize variances attributed to encoding, we used a repeated retrieval-encoding cycles procedure to bring initial learning to a memory accuracy criterion of 83% in all participants, so that the comparison of subsequent memory consolidation could be made with starting points as similar as possible.
We hypothesized small or no differences in short-delay memory consolidation between children and adults (Peiffer et al., 2020a; Wang et al., 2018), but less efficient long-delay consolidation in children in comparison to young adults, as memory consolidation over time is particularly dependent on the interactions between the HC and neocortical regions that are still maturing in preschool children (Ghetti & Bunge, 2012; Lebel et al., 2012; Shing et al., 2010a). Based on previous reports of maturational differences in hippocampal-cortical circuitry between preterm born and term born children (Nosarti & Froudist-Walsh, 2016), we predicted less efficient memory consolidation in preterm born children in comparison to term born children and young adults. We furthermore expected that thinner medial OFC and IFG would be associated with better short-delay memory consolidation, reflecting schema-integration and strategic control over memory, respectively (Badre & Wagner, 2007; Kharitonova et al., 2013; Leon et al., 2010; Shing et al., 2010b), and that larger HC volumes would be associated with long-delay memory consolidation (Gilboa, 2004; Østby et al., 2012b; Steinvorth et al., 2005; Tompary & Davachi, 2017). We expected these neural-behavioural associations to be more pronounced in children compared to adults. While our focus is on memory consolidation, we also explored potential group differences during the initial learning phase. Here, we expected similar final learning performance but less efficient learning efficiency of object-location associations in preterm born children in comparison to term born children. We also expected that thinner OFC would be related to more efficient final learning performance (Botdorf & Riggins, 2018; Burzynska et al., 2012; Østby et al., 2012a; Shing et al., 2010b).
2 METHODS
2.1 Participants
Preterm born children (born before 34 weeks of gestation) were recruited, either from the existing well-characterized cohort at the Department of Neurology and Neonatology of the Center for Chronically Sick Children (Sozialpädiatrisches Zentrum), Charité – Universitätsmedizin Berlin, or through word-of-mouth. For the recruitment of term born children, 2000 general research invitation letters were sent to randomly selected families with 6-year-old children in Berlin, of whom 60 families expressed interest in participation. After screening, 44 typically developing term born children were recruited to participate in the study. 35 term born young adults were recruited to participate in the study through advertisement in the newspaper, on the university campus, and through word-of-mouth.
All participants had normal vision with or without correction, no history of psychological or neurological disorders or head trauma and had a normal cognitive ability with an IQ > 85 assessed using the K-ABC II (Kaufman & Kaufman, 2015). Exclusion criteria were severe perinatal incidents such as hypoxia-ischemia, intraventricular hemorrhage or infection, necrotic enterocolitis, and/or severe bronchopulmonary dysplasia. 14 term born children were excluded due to incomplete task execution and missing data (n=7) or technical issues during data acquisition (n=7). 10 preterm born participants were excluded due to incomplete task execution and missing data (n=8). Four young adult participants were excluded due to incomplete task execution and missing data (n=2), and identification as an outlier (n=2) due to chance performance. Thus, the final sample size consisted of 30 typically developing term born children (12 female, mean age 6.37 years, SD = 0.36), 15 preterm born children (7 female, mean age 6.25 years, SD = 0.61), and 33 term born young adults (15 female, mean age 25.45 years, SD = 2.80; see Table 1).
Sample characteristics by age group (preterm born children, term born children, young adults) showing comparability between groups
All participants or their legal guardians gave written informed consent prior to participation. The study was approved by the ethics committee of the Goethe University Frankfurt am Main (approval E 145/18). The participants were compensated for participation on the three days of the experiment with 100 Euro.
2.2 Task design and procedure
2.2.1 Stimulus material and task
The stimuli were chosen based on the curriculum in social studies and science for the first and second grade of the German primary school (see similar procedure in Brod & Shing, 2019). Sixty different semantic themes (e.g., forest, farm, etc.) were chosen according to the ratings provided by four primary school teachers that assessed the familiarity of first graders with the topics. For each semantic theme, four scene pictures were combined with four thematically congruent object pictures resulting in four unique object-location associations (see Fig. 1 for example). We identified 18 possible placement areas, one of which was assigned to each object-location association (for more detailed information on task description refer to Fig. 1 and to the Methods section in the Supplementary Materials). We presented the task using Psychtoolbox-3 (Kleiner et al., 2007) software in Matlab 9.5, R2018b (The MathWorks, Inc., MA, USA).

(A) Initial Encoding Phase. For a set of 60 object-location pairs, participants saw the object followed by the same object superimposed on the scene at a particular location. Participants were instructed to remember 60 object-location pairs in total, memorizing the exact location of the object within the scene by creating a story or making a “mental photo” of the scene as the such semantic organization strategies aid the recollection of the information (Craik & Lockhart, 1972); (B) Learning Phase. Participants learned the correct location of the object within the scene during retrieval-encoding cycles. They had to choose the correct location out of three choices and received feedback for their response. After the feedback, the correct object-location associations were shown again. The cycles ended when participants provided correct responses to 83% of the trials (minimum two cycles, maximum four cycles). (C) Retrieval Phase. Participants had to choose the correct location of the object in the scene out of three options without feedback. Note: The retrieval phase was carried out inside the MRI scanner with a functional sequence, of which the data is not included as we focus on characterizing the retention rate behaviourally, both in terms of group comparison and relations to structural integrity.
2.2.2 Assessment of demographic and cognitive covariates
In addition to the experimental paradigm (memory consolidation task described below), IQ scores were assessed using the German version of the “Kaufman Assessment Battery for Children – Second Edition” (K-ABC II; Kaufman & Kaufman, 2015) in children and the “Wechsler Adult Intelligence Scale – Fourth Edition” (WAIS-IV; Wechsler, 2012) in young adults. General sociodemographic questionnaires to assess socio-demographic characteristics of the participants were applied as well. Other cognitive covariate tasks were also assessed, such as cognitive switching and object-location memory with immediate test but were not included in the current paper to retain the focus on memory consolidation (see Fig. 2).

The testing took place across three days. (i) At Day 0 participants had to learn 60 object-location associations. The experimental task started with the initial encoding of the first 30 object-location associations. The initial encoding was followed by a brief distraction task in which participants listened to and had to recall the string of numbers. The distraction task was followed by the learning phase with retrieval-encoding cycles until the 83% criterion. After a short break, the same procedure was repeated with the other half of 30 object-location associations. After the experimental task, several subtests of the K-ABC II Test *(e.g., Atlantis, Rover, Rebus, Riddle and Atlantis delayed) were administered to children, while young adults were tested with the WAIS-IV Test. (ii) On Day 1 (short delay) the retrieval was conducted. Participants had to retrieve 30 object-scene associations learnt on Day 0. In addition, children performed several subtests of the K-ABC II Test *(e.g., Expressive Vocabulary, Triangles, Pattern Reasoning). (iii) On Day 14 (long delay) the retrieval was conducted. Participants had to retrieve another 30 object-scene associations learnt on Day 0. In addition, children performed several subtests of the K-ABC II Test *(e.g., Patterns, Verbal Knowledge, Word Order). Note: * denotes specific subtests of K-ABC II distributed across three testing days.
2.2.3 Experimental procedure
Testing took place across three days. At Day 0, the experimental procedure began with a short training to familiarize participants with the task. The experimental task started with the initial encoding, which was followed by retrieval-encoding cycles until the 83% accuracy criterion was reached. On Day 1 (short delay) and Day 14, retrieval was conducted (see Fig. 2 for a detailed description of the experimental procedure).
2.3 Magnetic resonance imaging
MR images were acquired on a 3-T SIEMENS PRISMA scanner (Siemens Medical Solutions, Erlangen, Germany) using a 64-channel head coil. A MPRAGE (magnetization prepared rapid gradient echo) T1-weighted sequence was applied with the following parameters: time repetition/time echo/time to inversion/Flip Angle = 2400 ms/2.22 ms/1000 ms/8°, matrix 256 x 256, field of view = 256. Each scan took 6 min 38 s. Each volume consisted of 320 sagittal slices with voxel sizes 0.8 x 0.8 x 0.8 mm.
2.4 Data Analysis
2.4.1 MRI data processing
Volumes and cortical thickness measures used in the current analyses were derived using the anatomical pipeline of fMRIprep (version 20.2.1; Esteban et al., 2019), based on Nipype 1.5.1 (Gorgolewski et al., 2011). Brain tissue segmentation of cerebrospinal fluid, white-matter, and grey-matter was performed on the brain-extracted T1w using FAST (FSL 5.0.9; Zhang et al., 2001). Brain surfaces were reconstructed using recon-all (FreeSurfer 6.0.1; Dale et al., 1999). Volume-based spatial normalization to two standard spaces was performed through nonlinear registration with antsRegistration (ANTs 2.3.3), using brain-extracted versions of both T1w reference and the T1w template. Intracranial volume (ICV) was derived by the surfaced-based measures. Thickness measures were calculated from the distance between the reconstructed white matter and grey matter surfaces as well as from the reconstructed grey matter surface and cerebrospinal fluid boundaries (for a more detailed description of the segmentation procedure, please refer to the Methods section in Supplementary Materials).
2.4.2 Behavioural Data Analysis
The analyses of all behavioural measures were performed with R packages (Version 4.0.4, R Core Team, 2021) in R Studio 1.4.1106 (RStudio, Inc.). Throughout the analyses, the a levels were set to α < .05. We conducted a linear mixed-effect model for memory measures (accuracy defined as percentage of correct responses) using the lmer function from the lme4 package in R (Bates et al., 2015) and lmerTest (Kuznetsova et al., 2017). The linear mixed effect model was calculated with maximum-likelihood estimation and Subject as random intercept to account for between-subject variability in memory accuracy. As fixed factors, we included the within-subject factor of Session (final performance on Day0, short delay, and long delay) and the between-subject factor of Group (PC, TC, and YA). In addition, IQ, Sex, and Handedness were added as covariates into the model. The main effects were followed up with Tukey post-hoc multiple comparisons (Haynes, 2013). For group differences in learning measures, we conducted oneway independent analysis of variance (ANOVA). In case of violated assumptions of homogeneity of variances, a Games-Howell test was performed (Lee & Lee, 2018). Moreover, Games-Howell is especially suitable for the groups with unequal sample sizes. The effect size estimation was performed with omega squared (ω2) as a less biased estimate for reporting practical significance of observed effects (Finch & French, 2012; Okada, 2013; Troncoso Skidmore & Thompson, 2013). To determine the amount of variance explained by the model, we used partR2 package in R (Stoffel et al., 2020) with bootstrapping to calculate confidence intervals.
2.4.3 Analysis brain-behavioural associations
Based on the literature, we focused on the following brain regions of interest: HC volume, IFG cortical thickness (consisting of pars opercularis and pars triangularis), and medial OFC cortical thickness (Dudai, 2004; Kharitonova et al., 2013; Leon et al., 2010; Moscovitch & Gilboa, 2021; Østby et al., 2012b; Petrides, 2007). To control for head size, hippocampal volumes were adjusted for ICV using a residual approach, which has been suggested to be less susceptible to bias (Bathelt et al., 2018; Mathalon et al., 1993; Sanfilipo et al., 2004; Voevodskaya et al., 2014). Cortical thickness was adjusted for sex but not head size because cortical thickness and head size are not associated (Barnes et al., 2010; Mills et al., 2016). We conducted a linear mixed-effect model analysis of memory measures (accuracy defined as percentage of correct responses) using the lmer function from the lme4 package in R (Bates et al., 2015) and lmerTest (Kuznetsova et al., 2017). The linear mixed effect models were calculated with maximumlikelihood estimation and Subject as a random intercept. As fixed factors, we included the within-subject factor of Session (final performance on Day0, short delay, and long delay), the between-subject factor of Group ((PC, TC, and YA) or (children and adults)), and structural measures (HC volume, IFG cortical thickness and medial OFC cortical thickness) separately for left and right brain hemisphere, expecting task-related lateralization (Bauer et al., 2019; Ezzati et al., 2016; Hopf et al., 2013). In addition, Sex was added as a covariate in the model to account for possible differences in brain volumes and cortical thicknesses (Mills et al., 2016). Regression analyses were performed to assess associations between final learning performance and brain structural measures (medial OFC cortical thickness). Group differences in brain structure were assessed using one-way independent ANOVA, which was followed by either post hoc Tukey comparisons or a Games-Howell test, depending on the homogeneity of variances.
3 RESULTS
3.1 Final Learning and Learning Efficiency at Day 0
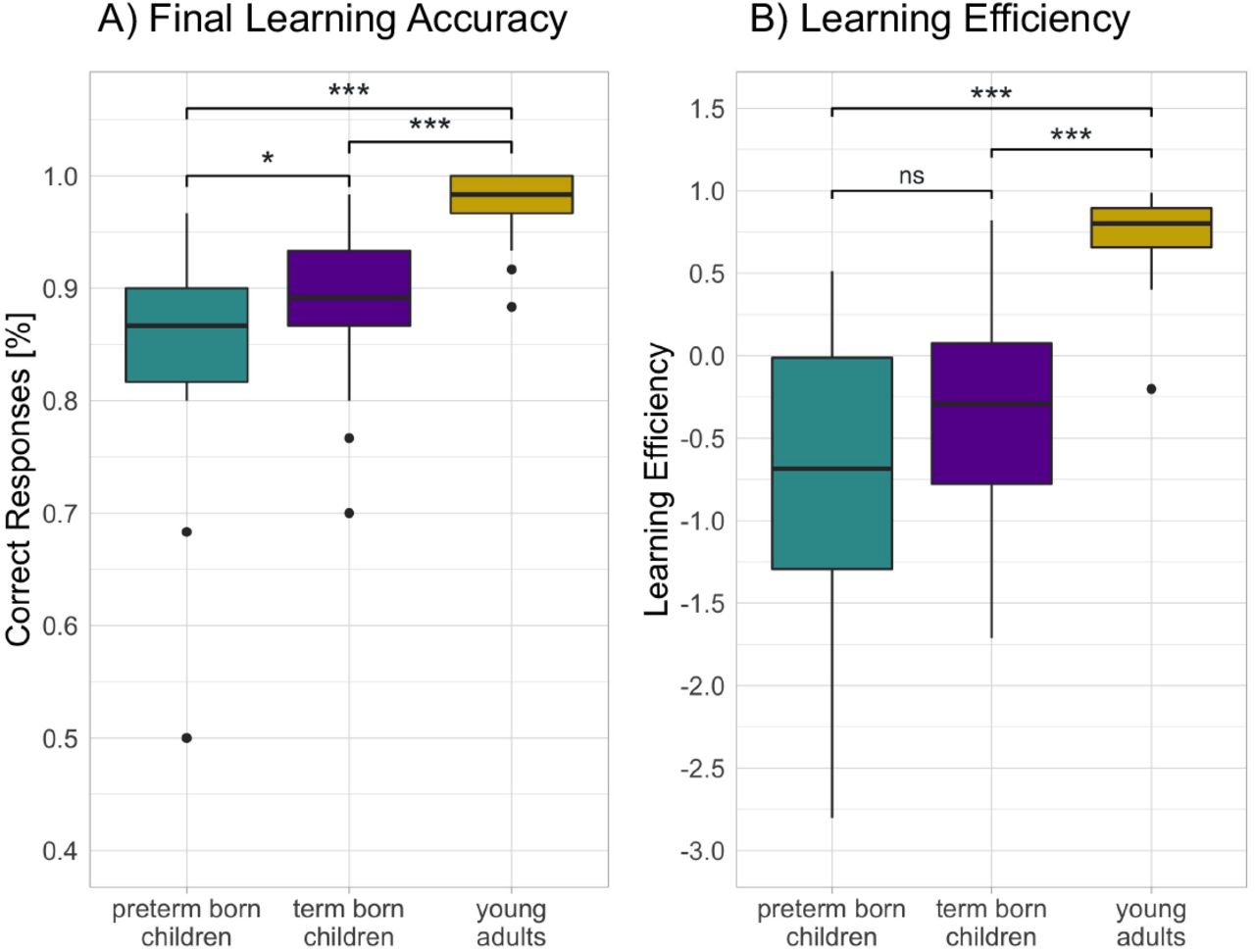
In the following, we first tested for potential group-related differences in learning object-location associations during the learning phase. The one-way independent analysis of variance (ANOVA) revealed a significant effect of Group on final learning accuracy, F(2,75) = 45.73, p < 001, ω2 = .53. Post hoc Games-Howell tests revealed that term born children learnt significantly more objectlocation associations than preterm born children, p = .049. Young adults showed significantly higher final learning accuracy than term born children, p < .0001, and preterm born children, p < .0001 (see Fig. 3). We also derived an index of learning efficiency, which is calculated as the average of the standardized z scores of initial encoding, the learning slope, and final learning accuracy (Zerr et al., 2018). There was a significant effect of Group on learning efficiency, F(2,75) = 48.19, p < 0.001, ω2 = .55. Post hoc Games-Howell tests revealed that learning efficiency did not differ between preterm born and term born children, p = .27, while the learning efficiency of young adults was significantly higher than that of term born children, p < .0001, and that of preterm born children, p < .0001 (see Fig. 3).

(A) Final learning accuracy shows the percentage of correct responses after learning was completed in preterm born children, term born children and young adults. (B) Learning efficiency implies the learning rate and immediate retention rate for object-location associations as introduced by Zerr et al (2018). Error bars indicate Q1/3 ± 1.5·IQR; *p < .05; **p < .01; ***p < .001(significant difference); ns: non-significant difference.
3.2 Memory Consolidation
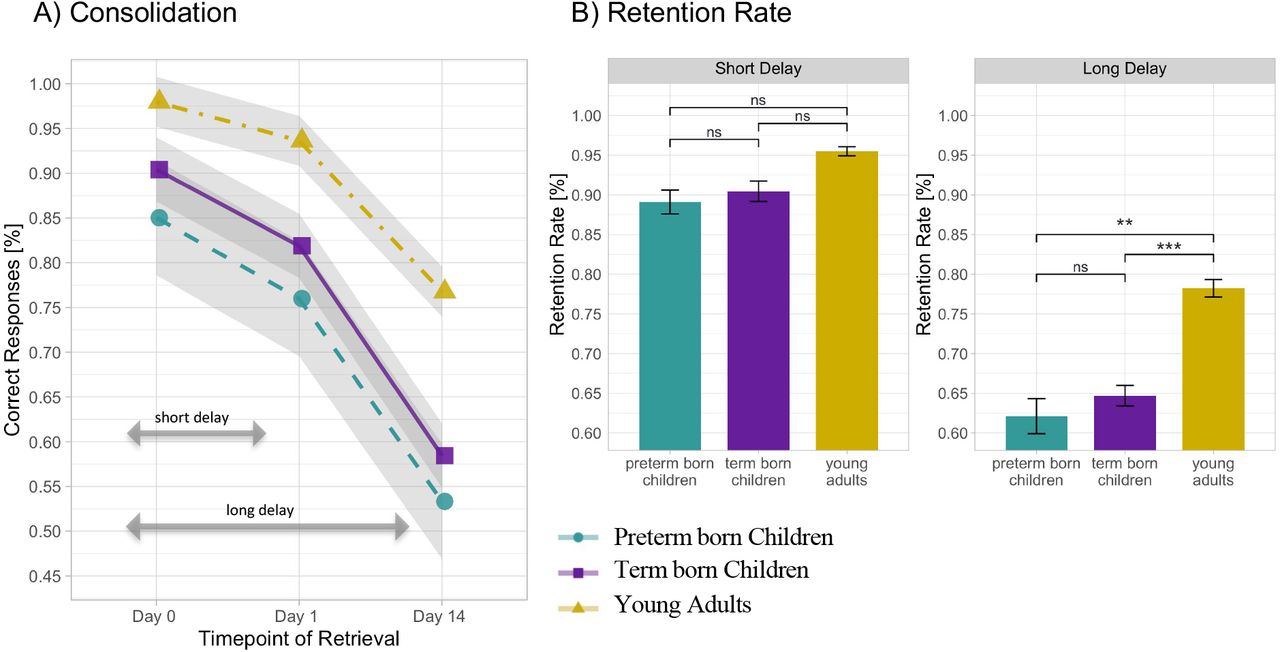
In the following, group differences in short- and long-delay memory consolidation are presented. The linear mixed-effects model for retrieval accuracies of learned object-location pairs explained a significant amount of variance, R2 = .69, 95% CI [.66, .76] (for a full overview, please see Table 2 in Supplementary Material). We observed a significant Session x Group interaction, F(4,156) = 4.71, p = .001, ω2 = .08. As a follow-up to this interaction, we calculated model-based Tukey post hoc comparisons for short and long delay separately. The between group comparisons for short delay revealed that (i) retrieval accuracy in all groups after one night of sleep declined significantly (all p <.05); (ii) there was no difference in the slope of accuracy decline between groups (all p > .12), indicating a comparable efficiency of short-delay memory consolidation among 6-year-old children, irrespective of birth status, and young adults. For the long delay retrieval after a period of two weeks, Tukey post hoc between group comparisons revealed (i) a significant decline in long delay retention rate in all groups (all p < .0001); (ii) no difference in long delay retention rates between preterm born and term born children, b = .003, t(162)= .08 p = .94; (iii) significantly higher long-delay retention rates in young adults than in both preterm born children, b = .105, t(162) = 3.14, p = .002, and term born children, b = .108, t(162) = 3.97 p = .0001, indicating more efficient long-term memory consolidation in young adults in comparison to 6-year-old children, irrespective of their birth status (see Fig. 4).
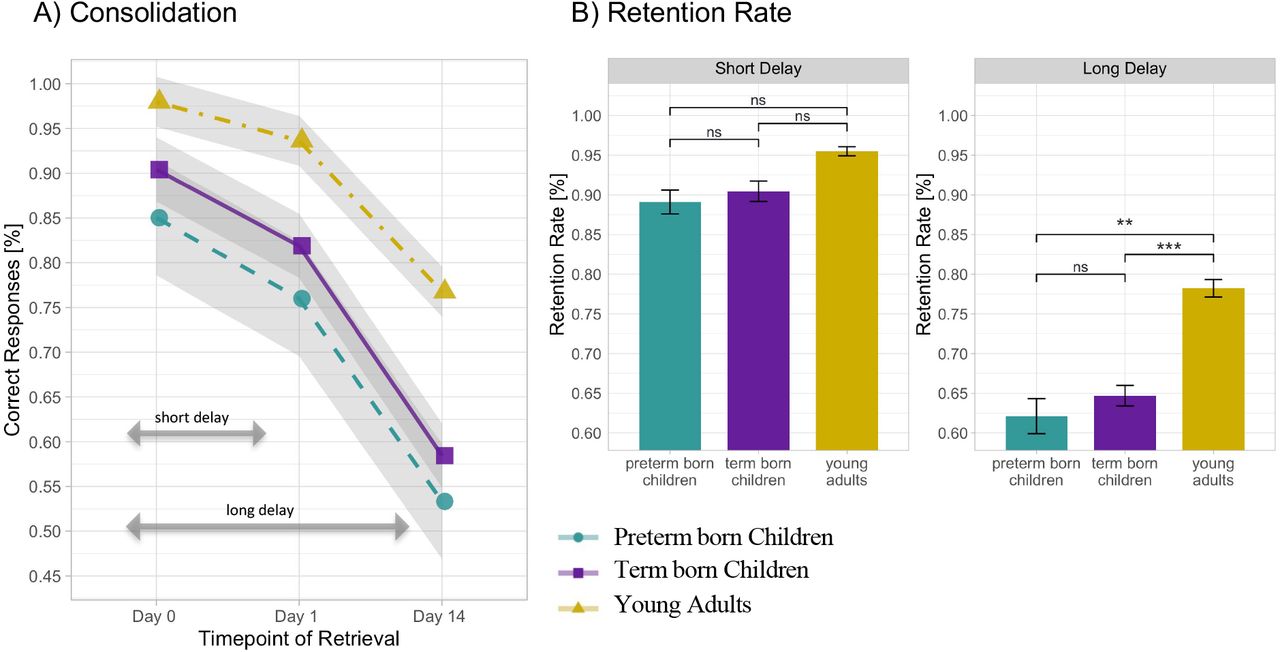
(A) Memory consolidation over the course of two weeks operationalized by percentage of correctly retrieved objectlocation associations learned on day 0 (final accuracy after learning), on day 1 (after one night of sleep) and on day 14 (after two weeks). Shaded areas indicate 1 standard error of the mean. (B) Retention rate for short (after one night of sleep) and long (after two weeks) delay across preterm born children, term born children and young adults. Error bars indicate 1 standard error of the mean. *p < .05. **p < .01. ***p < .001(significant difference), ns: non-significant difference. P-values use Tuckey correction for multiple comparisons.
Results of the analyses on group differences for brain structural measures (HC volumes, medial OFC thickness, IFG thickness). Statistical values are shown for the ANOVA analyses and follow-up post hoc tests.
3.3 Brain-behavioural relationships
3.3.1 Hippocampal Volume and Memory Consolidation
The between group analyses showed that residualized left and right HC volumes were comparable between term born and preterm born children, while adults showed bigger left and right HC volumes in comparison to both groups of children (see Table 2). Based on this observation, term born and preterm born children were combined into one group for the subsequent analyses (referred to as the children group). Regarding memory consolidation, separate linear mixed-effects models were conducted with left and right HC volumes as predictors of memory retention rate, respectively. Only right HC volumes, but not left HC, significantly predicted retention rates, irrespective of short or long delay and age group, R2 = .70 95% CI [.66, .77], F(1,75) = 5.98, p = .016, ω2 = .06 (see Fig. 5). Moreover, there was a significant right HCx Group interaction, F(1,75) = 6.01, p = .016, ω2 = .06, indicating an age effect in the relation of right HC and memory consolidation. This interaction was probed by conducting two separate multilevel models for the group of children and the group of young adults with the same setup as above but without the Group factor, as in (Field et al., 2012). The analysis showed that for children, higher right HC volume was significantly associated with long-delay retention rates, b = .17, t(2.11), p = .037, but not short-delay retention rates, b = .08, t(1.04), p = .29, while such association was not significant in adults (all p > .65). An overview comprising the statistics of all effects of the applied models can be found in the supplementary materials (see Supplementary Results, Tab. S4-S7).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Scatterplots depicting associations between short-delay and long-delay retention rates with residualized right HC volume. Retention rate on the y-axis is measured as a proportion between final accuracy on Day 0 and memory performance on Day 1 (short delayday1/day0) and between final accuracy on Day 0 and memory performance on Day 14 (long delayday14/day0). On the x-axis the residualized right HC volume was corrected for head size (using ICV) using the residual approach (Mathalon et al., 1993; Voevodskaya et al., 2014). (B) Scatterplot showing the relation between the final learning accuracy on Day 0 and right medial orbitofrontal cortical thickness (centered). HC: Hippocampus; ICV: intracranial volume.
3.3.2 Frontal Cortical Thickness and Memory Consolidation
The between group analyses showed that left and right medial OFC was thicker in preterm born children in comparison term born children, while adults showed thinner right medial OFC than both child groups. Furthermore, the between group analyses showed that left and right IFG cortical thickness (i.e., pars opercularis and pars triangularis) were comparable between term born and preterm born children, while adults had thinner left and right IFG cortical thickness in comparison to both child groups. As we expect a similar relation of cortical thickness measures to memory measures in both groups of children, they were combined into one group for the subsequent analyses (referred to as the child group; Nam et al., 2015).
With respect to memory consolidation, linear mixed-effects models were conducted separately with left and right medial OFC thickness as predictors of memory retention rate. Only a main effect of right medial OFC thickness, F(1,75) = 4.64, p = .03, ω2 = .05, was observed, indicating that thinner right medial OFC is related to higher overall retention rates irrespective of age group and delay. No such effect was observed for the left medial OFC (all p > .09). The linear mixed-effect models conducted with left and right IFG thickness as predictors of memory retention rate revealed no significant effects (all p > .19).
3.3.3 Exploratory analysis with final learning performance
Finally, we also explored brain-behavioural relationships for day 0 final learning performance. First, the association between the medial OFC thickness and final learning performance was investigated. As revealed by regression analyses, there was a marginally significant association between right medial OFC thickness and final learning performance, F(1,71) = 3.47, p = .06, ω2 = .03, which differed by group, F(1,71) = 4.88, p = .03, ω2 = .05. Moreover, left OFC thickness was also significantly associated with final learning performance, F(1,71) = 9.43, p = .003, ω2 = .10, irrespective of age group, F(1,71) = 1.88 p = .06, ω2 = .02. Follow up tests on observed groups differences revealed that in children, thinner right medial OFC thickness was predictive of final learning performance while thinner left OFC was associated with final learning performance in both groups of children and in adults (see Fig. 5). For completeness, we also explored the relationship between the IFG and HC with final learning performance, which did not yield any significant results (all p > .26). Therefore, only the medial OFC showed significant relationship with final learning performance.
4 DISCUSSION
In the present study, we investigated memory consolidation of object-location associations after both short delay (after one night of sleep) and long delay (after two weeks), in preterm born and term born 6-year-old children as well as young adults, and examined their associations with medial temporal and frontal structural brain measures. Given postulated differences among the three groups in terms of cognitive functioning and structural brain integrity in general, we took several steps to hone in on differences in memory consolidation by reducing group differences in general intellectual functioning or in the encoding of new information. All participants learned the object-location associations to a high degree of accuracy at day 0. Nevertheless, we found that preterm born children showed impaired final performance in learning object-locations associations when compared to term born children, who in turn were outperformed by young adults. We therefore took performance on day 0 into account in our statistical analysis of memory consolidation. Here, our main findings are: (i) preterm born children, term born children, and young adults showed comparable short-delay retention rates after one night of sleep; (ii) the two child groups showed comparable retention rates over long delay, but these rates were significantly lower than those of young adults. In other words, children showed steeper forgetting of information over a longer time period.
Taking a step further, we tested for relationships between structural measures of key brain regions involved in memory consolidation and behavioural performance. In both children and young adults, a thinner medial orbitofrontal cortex correlated with higher retention rates, while no such association was found for IFG. Furthermore, we found that memory retention rates were positively associated with larger right HC volume uniquely in children. In addition, thinner medial OFC was associated with higher final learning performance in both children and adults. These results will be discussed in the following sections.
4.1 Short- and long-delay memory consolidation
4.1.1 Similar short- and long-delay memory consolidation in term born and preterm born children
Contrary to our hypothesis, there was no difference in short-delay and long-delay retention rates between preterm born and term born children, indicating similar memory consolidation performance despite premature birth. This finding may be attributed to a highly selected cohort of preterm born children that was comparable to the term born children in their general ability to learn and acquire knowledge as well as exhibiting comparable world knowledge according to subscales of K-ABC II (see Supplementary Materials for further information). These abilities benefit long-delay memory consolidation that relies on schemas or complex knowledge systems (Aghayan Golkashani et al., 2021; Bartlett, 1995). Interestingly, it has been shown that comparable memory performance can be achieved in adult preterm born individuals by engaging different brain circuits during encoding and successful retrieval (Brittain et al., 2014; Narberhaus et al., 2009; Salvan et al., 2014; Tseng et al., 2017). This may indicate a high degree of neuroplasticity in individuals born prematurely to already begin to compensate for alterations in the brain early in developmental, accounting for our findings of comparable memory consolidation between term born and preterm born children.
4.1.2 Similar short-delay but less efficient long-delay memory consolidation in term born and preterm born children in comparison to young adults
As expected, adults and children showed comparable short-delay retention rates, indicating that short-delay memory consolidation after one night of sleep in 6-year-old children was as efficient as in young adults irrespective of preterm born status. However, both groups of children showed steeper accuracy percentage change and thus lower long-delay retention rates in comparison to young adults, indicating reduced efficiency in retaining memory of complex associative information over longer time periods in children. This evidence is in line with the findings on comparable or even more efficient short-delay memory retention rates in primary school children than in young adults (Peiffer et al., 2020; Wang et al., 2018), which may be due to a stronger effect of overnight sleep on short-delay memory consolidation in children in comparison to young adults, compensating for possible maturity-related mnemonic benefits in adults and also pointing at an early advantage of sleep for preschool children (Peiffer et al., 2020). On the other hand, superior long-delay memory consolidation in young adults may be attributed to their more extensive world knowledge and therefore more elaborate schemas that may be especially important for ensuring better accessible memory representations in long delay over short delay (Craik & Lockhart, 1972; van Kesteren et al., 2012). Moreover, these mnemonic benefits may be augmented by mature HC-vmPFC interaction that underlies long-term memory consolidation in young adults (Takashima et al., 2006; Yamashita et al., 2009). In children, the extended developmental trajectory into mid-childhood for the HC and into adulthood for the vmPFC (Gogtay et al., 2004b; Keresztes et al., 2017b; J. K. Lee et al., 2016b; Shing et al., 2010a) may have limited the efficiency of consolidating memory into neocortical regions over long delay.
4.1.3 Thinner right HC volume is associated with long-delay memory consolidation in children and thinner right mOFC thickness is associated with overall memory consolidation in all age groups
The premature birth-related brain structural differences previously reported in the literature (Nosarti et al., 2014b; Nosarti & Froudist-Walsh, 2016) could not be found in our study. Contrary to our expectations, there was no difference in HC volume or IFG cortical thickness between preterm born children and term born children. The only difference found within our ROIs was that preterm born children showed thicker medial OFC in comparison to term born children. Adults showed larger HC volumes and thinner IFG and medial OFC than both groups of children, which is in line with existing findings on developmental trajectories of brain morphology (see Hedman et al., 2012 for an overview).
Importantly, in children, higher right HC volumes were positively associated with long-delay memory retention, indicating that more mature right HC is associated with better memory consolidation over longer time periods in children irrespective of premature birth status. This finding is convergent with the established critical role of the HC in relational memory (Konkel, 2009). Concerning lateralization, in line with our findings, Hopf et al. (2013) could show that more adult-like right HC lateralization in children was associated with superior memory on a relational memory task. Moreover, activation in the right HC has been reported to be associated with spatial memory processing (Bohbot et al., 1998; Ezzati et al., 2016) and with memory for locations within an environment (Burgess et al., 2002). The lack of association between HC volumes and memory retention rates in young adults may be attributed to the relative independence of memory consolidation processes on HC volumetric characteristics in adults compared to children, in which volumetric increase corresponds to maturation. It has been proposed that in young adults, HC involvement in memory consolidation is more pronounced in the functional dynamic interplay with other neocortical regions (Frankland & Bontempi, 2005).
On the other hand, thinner right medial OFC was related to higher overall retention rates irrespective of age group, in line with its role in schema-related memory integration through perceived congruency between to-be-learnt information and prior knowledge (Brod & Shing, 2018; Tse et al., 2007). This finding goes beyond previous functional evidence, showing that a structurally thinner medial OFC is also related to better memory formation and retention over time. Convergingly, Bauer et al. (2019) showed that right medial OFC thickness was related to memory of self-derived knowledge in 5 to 8 years old children. Memory recall has also been shown to be related to OFC thickness in 8 to 19 years old participants, irrespective of age (Østby et al., 2012a). Therefore, our results suggest that structural measures of OFC are related to memory consolidation as well, with thinner OFC indicating higher maturity and being associated with better memory integration with prior knowledge regardless of age (Schlichting & Preston, 2015).
Contrary to our expectations, IFG thickness was not associated with memory consolidation irrespective of age group and delay, despite functional brain imaging findings suggesting an association between the IFG and semantic organizational strategies for verbal learning (Savage et al., 2001) and monitoring strategic control processing over retrieval of memory (Badre & Wagner, 2007; Kuhl et al., 2012), as well as semantic elaborative strategies during memory retrieval that arise deliberately from prior encoding of distinctiveness (Han et al., 2012; Raposo et al., 2009). Therefore, structural morphology of IFG may not be associated with memory consolidation beyond its functional involvement in encoding and retrieval when the focus is not on distinctness or semantic elaborations aimed to capitalize on it for long-term memory retention.
4.2 Learning
4.2.1 No difference in learning efficiency between preterm and term born children but reduced learning final performance in preterm in comparison to term born children
While our focus was on memory consolidation, we also explored groups differences in Day0 learning. We found no difference between preterm born and term born children with respect to learning efficiency, indicating a comparable rate of learning. However, preterm born children showed reduced final learning accuracy in comparison to term born children. This finding may be related to the impaired visuo-spatial pattern recognition (Marlow et al., 2007; Schermann & Sedin, 2007) and visuo-spatial memory (Omizzolo et al., 2014b; Thompson et al., 2013b) reported for preterm born children. Indeed, in our sample, despite comparable general cognitive functioning, there was a small difference found in the simultaneous subscale of the K-ABC II Test (refer to Supplementary Materials for further information), suggesting a lower ability for simultaneous processing of visuospatial information in preterm born children, which may have limited the amount of information they could learn in our task.
4.2.2 Less efficient learning in term born and preterm born children in comparison to young adults
In line with our expectation, both groups of children were outperformed by young adults in final learning accuracy and learning efficiency. These findings indicate that young adults were able to retain more information after just a single exposure to the stimuli pairs, learn faster, and retrieve with higher final accuracy than both groups of children. The superior learning efficiency performance of young adults may be attributed to more efficient associative binding as well as better ability to exert strategic control over memory (Shing et al., 2008b, 2010b). The ongoing development of the PFC and HC in children as well as their lesser knowledge and poorer integration of newly learned information may have limited the efficiency of learning during the encoding-retrieval cycles (Shing et al., 2010c).
4.2.3 Thinner medial orbitofrontal cortex is related to more efficient final learning performance in children and adults
Final learning accuracy was positively related to thinner right medial OFC in children and to thinner left medial OFC across all participants. In line with this, a positive association between thinner frontal lobes with better immediate (30 minutes delay) memory was observed in visuospatial memory task in children aged 6 to 15 years (Sowell et al., 2001). In addition, Østby et al. (2012) showed that thinner left OFC was related to higher memory recall 30 minutes after learning independently of age in participants aged 8 to 19 years. Thus, these findings show that for to-be-remembered complex associations that are congruent with prior knowledge, thinner medial OFC is related to better memory formation (Brod & Shing, 2018). This association was already present bilaterally at the age of six, indicating more intense brain-behaviour interactions in the maturing brain, which continues to persist in mature brains in young adulthood as well.
5 LIMITATIONS
Several limitations of the current study should be noted. Despite our procedure of learning to the criterion to maximize comparability of retention rates, we nevertheless observed group differences in initial memory performance. Future studies may incorporate individualized itembased approach of learning to criteria, excluding correctly remembered items from further learning cycles to ensure faster learning and lessening the overall task workload (Karpicke & Roedigeriii, 2007; McDermott & Zerr, 2019; Zerr et al., 2018). Moreover, due to the small sample of preterm born children, we may have been underpowered to detect memory consolidation differences in term and preterm born children. Therefore, future studies with a larger preterm born sample would help to draw more robust conclusions about potential differences in memory consolidation due to premature birth status (or gestational age) in well-characterized cohorts. In addition, the current findings concentrate mainly on associative memory of schema-congruent information. Future studies should investigate how violations of knowledge, namely schema-incongruent information, may impact the learning of associative information and their subsequent consolidation in short and long delay memory.
Conflict of interest statement
We have no known conflict of interest to disclose.
Data availability statement
The datasets generated and analysed during the current study are available from the corresponding authors upon reasonable request.
Ethic approval statement
The study was approved by the ethics committee of the Goethe University Frankfurt am Main, Germany (Nr. E 145/18).
Author contributions
Y.L.S, C.B., A.K secured funding. I.S and Y.L.S, C.B., A.K contributed to conception and design of the study. I.S., H.S., N.W.-C., and P.L. performed data collection and data curation. I.S., P.L., and M.B performed the statistical analysis. IS wrote the first draft of the manuscript, P.L. and M.B. wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Acknowledgements
We thank all the children and parents who participated in the study. This project was funded by the Deutsche Forschungsgemeinschaft (DFG; German Research Foundation, Project-ID 327654276, SFB 1315, “Mechanisms and Disturbances in Memory Consolidation: from Synapses to Systems”). The work of YLS was also supported by the European Union (ERC-2018-StG-PIVOTAL-758898).
Footnotes
↵* Yee Lee Shing and Claudia Buss should be considered joint senior author.
7 REFERENCES




