

Abstract
Improving our understanding of Mayaro virus (MAYV) ecology is critical to guide surveillance and risk assessment. We conducted a PRISMA-adherent systematic review of the published and grey literature to identify potential arthropod vectors and non-human animal reservoirs of MAYV. We searched PubMed, Embase, Web of Science, SciELO and grey-literature sources including PAHO databases and dissertation repositories. Studies were included if they assessed MAYV virological/immunological measured occurrence in field-caught, domestic, or sentinel animals or in field-caught arthropods. We conducted an animal seroprevalence meta-analysis using a random effects model. We compiled granular georeferenced maps of non-human MAYV occurrence and graded the quality of the studies using a customized framework. Overall, 57 studies were eligible out of 1523 screened, published between the years 1961 and 2020. Seventeen studies reported MAYV positivity in wild mammals, birds, or reptiles and five studies reported MAYV positivity in domestic animals. MAYV positivity was reported in 12 orders of wild-caught vertebrates, most frequently in the orders Charadriiformes and Primate. Sixteen studies detected MAYV in wild-caught mosquito genera including Haemagogus, Aedes, Culex, Psorophora, Coquillettidia, and Sabethes. Vertebrate animals or arthropods with MAYV were detected in Brazil, Panama, Peru, French Guiana, Colombia, Trinidad, Venezuela, Argentina, and Paraguay. Among non-human vertebrates, the Primate order had the highest pooled prevalence (PP) at 13.1% (95% CI: 4.3-25.1%). From the three most studied primate genera we found the highest prevalence was in Alouatta (PP: 32.2%, 95% CI: 0.0-79.2%), followed by Callithrix (PP: 17.8%, 95% CI: 8.6-28.5%), and Cebus/Sapajus (PP: 3.7%, 95% CI: 0.0-11.1%). We further found that MAYV occurs in a wide range of vectors beyond Haemagogus spp. The quality of evidence behind these findings was variable and prompts calls for standardization of reporting of arbovirus occurrence. These findings support further risk emergence prediction, guide field surveillance efforts, and prompt further in-vivo studies to better define the ecological drivers of MAYV maintenance and potential for emergence.
Author Summary Mayaro virus (MAYV) is an emerging tropical public health threat in the Americas. We conducted a georeferenced, quality-graded systematic review to evaluate the current evidence regarding MAYV occurrence in non-human vertebrates and arthropods. Overall, 57 studies were eligible out of 1523 screened, published between the years 1961 and 2020. Seventeen studies reported MAYV positivity in wild mammals, birds, or reptiles and five studies reported MAYV positivity in domestic animals. MAYV positivity was reported in 12 orders of wild-caught vertebrates, most frequently in the orders Charadriiformes and Primate. Our systematic review identified 12 orders of wild-caught vertebrates and seven mosquito genera with evidence of MAYV occurrence. Primates had the highest pooled MAYV prevalence according to a seroprevalence meta-analysis. The graded quality of evidence behind these findings was variable and prompts calls for standardization of reporting of MAYV and perhaps other emerging arbovirus occurrence in animals and vectors. This study provides important information for public health authorities and disease ecologists concerned with the growing threat of MAYV in Latin America. Our analysis provides a foundation for future laboratory and field studies focused on the MAYV transmission cycle.
Introduction
First detected in Trinidad in 1954 [1], Mayaro virus (MAYV) is a zoonotic Alphavirus that is endemic in several Latin American countries. Like Chikungunya virus (CHIKV), MAYV may cause complications such as debilitating arthralgia but often presents with a non-specific constellation of symptoms and signs that may be clinically indistinguishable from other vector borne diseases such as dengue or Zika [2]. There is no current licensed vaccine or antiviral treatment for MAYV infections, and the current standard of clinical treatment is supportive care only [2, 3].
MAYV has caused periodic outbreaks in humans in Brazil [4, 5], Bolivia [6], and Venezuela [7], while surveillance studies and serological surveys have detected MAYV in humans in several countries throughout the Americas including Peru [8], Suriname [9], Mexico [10], Colombia [11], French Guiana [12], and Haiti [13]. These findings demonstrate widespread circulation of the virus throughout the region. A recent 2019 epidemiological alert by the Pan American Health Association (PAHO) has emphasized the need for increased awareness of and extended surveillance for this emerging virus in the Americas [3]. However, the precise areas of risk from MAYV throughout the Americas remain unclear. Understanding the ecology and distribution of MAYV remains a major obstacle in predicting areas that are at high risk of transmission to humans and domestic animals.
Current evidence suggests that MAYV is maintained in nature through a sylvatic transmission cycle involving mosquito vectors and non-human animal reservoirs. Therefore, human MAYV cases reported to date likely represent direct sylvatic spillovers. Residing near forested areas [12] and hunting in the rainforest [14] have been identified as risk factors for MAYV infection in humans, highlighting the importance of the sylvatic transmission cycle and the potential for spillover events.
Identification of the non-human vertebrate animals (i.e., reservoirs) involved in MAYV transmission is an important step in delineating the human populations at greatest risk. The spillover of MAYV into humans represents a complex interaction of processes involving the density and distribution of reservoirs and vectors, as well as the prevalence and intensity of infection among reservoirs [15].
Identifying the non-human to, the challenges associated with establishing evidence of infection in wild animal populavertebrates that may serve as MAYV reservoirs is a difficult task due to a myriad of issues including, but not limited tions [16, 17]. High seroprevalence of a pathogen in an animal population does not necessarily implicate a given host as an efficient reservoir; conversely, low seroprevalence at a single point in time cannot definitively rule out an animal as a reservoir [17]. Due to the relatively short viremia of MAYV (approximately 3-10 days) molecular assays may be unsuccessful in detecting virus [18], necessitating the use of serological assays such as hemagglutination-inhibition (HI) assays, enzyme-linked immunosorbent assays (ELISA), or plaque-reduction neutralization tests (NT).
Several studies have been conducted to clarify the precise vertebrate hosts that may serve as MAYV reservoirs. High seroprevalence among non-human primates (NHPs) in Brazil [19], French Guiana [12], and Panama [20] provides evidence that NHPs may play an important role in the MAYV transmission cycle. MAYV antibodies have also been detected in mammals including rodents and marsupials [21] as well as several avian species [19]. Unfortunately, there is significant heterogeneity in the study methods used to identify potential MAYV reservoirs and there remains a high level of uncertainty surrounding the role of various non-human vertebrate species in the MAYV transmission cycle.
Studies have also been conducted in wild-caught mosquito populations as well as in controlled laboratory conditions in order to identify potential arthropod vectors of MAYV. One study in Brazil [19] suggested that the canopy-dwelling Haemagogus janthinomys mosquito is an important vector of MAYV. Additional mosquito species including Aedes aegypti, Ae. albopictus, and several anopheline species have been shown to be competent vectors in laboratory settings [22-24], posing a potential but as yet theoretical risk of urban MAYV cycles. The occurrence of MAYV in the city of Manaus has also led to concerns about the involvement of Aedes mosquitoes in a MAYV urban transmission cycle [25].
Although many non-human vertebrate animals and arthropod species have been proposed as capable MAYV reservoirs or vectors, our understanding of the MAYV transmission cycle and ecology remains limited. Collating and evaluating the current evidence regarding the potential MAYV reservoirs and vectors are important steps in characterizing MAYV transmission ecology and identifying the communities at greatest risk for MAYV outbreaks. Therefore, the goal of this systematic review is to evaluate the current evidence regarding MAYV occurrence in non-human vertebrates and arthropods. We present here the first structured evaluation of the potential vector and non-human reservoir range of MAYV, including the development of custom criteria for grading the quality of evidence of arbovirus occurrence in invertebrate and vertebrate non-human hosts.
Methods
This systematic review and meta-analysis were conducted according to the PRISMA 2020 Checklist [26] (see S1 Table). A protocol was developed but was not uploaded to PROSPERO.
Information Sources
We conducted a systematic review of original research articles, reports, and dissertations that attempted to identify potential non-human animal reservoirs or arthropod vectors of MAYV. We first searched Embase, Web of Science, PubMed, and SciELO databases for English, Spanish, and Portuguese language articles published between 1954 (the year MAYV was first isolated) and March 21, 2020. We searched all databases using the highly sensitive search term “Mayaro”. A PubMed alert using the search term “Mayaro” was also set to capture any additional studies that were published between the initial search and May 2021. This database search was extended using bioRxiv (https://www.biorxiv.org/) and medRxiv (https://www.medrxiv.org/) pre-print databases. We complemented these database search results with ‘grey literature,’ including hand-searched bibliographies of MAYV review articles (including systematic reviews), dissertations from several Brazilian university repositories, the Pan American Health Organization (PAHO) Institutional Repository for Information Sharing database (iris.paho.org), the GIDEON database (https://www.gideononline.com/), and GenBank [27] (https://www.ncbi.nlm.nih.gov/genbank/). In addition, we searched conference handbooks that are available online (2004-2019) from the American Society of Tropical Medicine and Hygiene (https://www.astmh.org/annual-meeting/past-meetings).
Eligibility Criteria
We included studies that evaluated past or current MAYV infection in non-human vertebrates using methods including virus isolation, molecular detection, and serosurveys. We also included studies that screened arthropods for MAYV using virus isolation and molecular detection. Original research studies were considered for eligibility if they assessed MAYV positivity in field-caught, captive, or sentinel non-human vertebrates or field-caught arthropods. Studies that met any of the following exclusion criteria were not included: studies involving only humans; studies not reporting original data (e.g., review articles, perspective pieces, editorials, recommendations, and guidelines); duplicate studies; in vitro studies such as vector cell-line or mammal cell line experiments; laboratory-based vector competence studies that did not explicitly demonstrate the detection of MAYV in a wild-caught vector; in-vivo lab-reared animal studies or any laboratory-based study that experimentally inoculated an animal to test theoretical reservoir status.
Selection process
All articles were organized using EndNote software version X9 (Clarivate, Philadelphia, Pennsylvania, USA), and data were abstracted into a Microsoft Excel table. Two reviewers independently screened all titles and abstracts to determine articles that could immediately be excluded and articles that should be included in the second stage of review. Results were compared to reconcile any differences between the two reviewers. The first and second reviewers then independently read the full text of potentially eligible articles identified through screening and selected the articles that were candidates for inclusion in the study. Results were compared to reconcile any differences between the two reviewers. A third-party reviewer adjudicated when consensus was not reached between the two reviewers during the first or second stage review. From those studies deemed eligible, data were extracted from articles by one reviewer using the data abstraction tool in Microsoft Excel.
Data abstraction
Relevant information was abstracted by one reviewer in an Excel sheet. Information for each article was abstracted across several domains including publication details (author and affiliation, study title, study funding), study methods (date and location of study, study design, laboratory methods to assess MAYV positivity), and study results (sample size, taxonomic classification, proportion of animals testing positive for MAYV, location of vertebrates/arthropods testing positive for MAYV). A second reviewer randomly selected and reviewed five articles for review to validate the data abstraction process.
Grading quality of evidence
We developed a customized grading system to assess the quality of each study included in our review. Several published studies have employed a similar grading system to assess evidence quality of included articles [28-30]. We assigned each study in our systematic review a grade for each of four quality items: clarity of research question/objective (Was the research question/objective clearly described and stated?); description of study methods (Were the study methods presented in a reproducible way?); description of sampling methods (Was the sampling method described in detail?); and validity of diagnostic tests (Was MAYV positivity measured in a valid way?). For each quality item, eligible studies were assigned a score of 3 (strong evidence), 2 (moderate evidence), 1 (weak evidence), or unable to judge. Studies were deemed unable to judge if the information provided was insufficient to assign quality scores (e.g., a single GenBank entry or conference abstract).
A score of 3 was assigned for the description of sampling methods item if authors thoroughly described the type of trap used, the habitats in which traps were set, how often traps were checked, and the results of trapping (i.e., were animals reported to the species level). For studies that assessed MAYV in vertebrate animals, a score of 3 was assigned for the validity of diagnostic tests item if MAYV positivity was assessed using RT-PCR, viral culture, or high-specificity serological method (i.e., plaque reduction NT); a score of 2 was assigned if MAYV positivity was assessed using non-specific serological assay (i.e., HI and ELISA); and a score of 1 was assigned if MAYV positivity was based on presumptive exposure only with no confirmatory assay. For studies that assessed MAYV in arthropods, a score of 3 was assigned for this item if MAYV positivity was assessed using viral culture; a score of 2 was assigned if MAYV positivity was assessed using RT-PCR or metagenomics; and a score of 1 was assigned if MAYV positivity was based on presumptive exposure only with no confirmatory assay. A score of “NA” was assigned for the validity of diagnostic tests item if studies did not detect MAYV positivity in any animal or arthropod samples.
Quality review scores were recorded in two different Excel documents for animal reservoir studies and arthropod vector studies, respectively. Two reviewers independently graded the evidence quality for each study and results were compared to reconcile any differences between the two reviewers. A third-party reviewer adjudicated if consensus was not reached between the two reviewers.
Data analysis
Descriptive Analysis
Descriptive statistics were presented by species for potential animal reservoirs showing the total sample size, proportion infected, and locations of infected animals. Descriptive statistics were presented by species for potential arthropod vectors showing the total sample size and total pools tested for virus (if applicable), the number of MAYV isolates or PCR-positive pools, and locations of infected arthropods. Maps were developed using ArcGIS software [31] to display the geographic distribution of MAYV-positive animals and vectors.
Pooled Analysis
Due to the heterogeneity of study designs and outcome measurements, a quantitative meta-analysis across all eligible studies was not possible. Instead, we conducted a seroprevalence meta-analysis using the studies that reported MAYV seroprevalence (i.e., using serological methods including HI, ELISA, or NT) in non-human vertebrate animals. Pooled prevalence estimates were stratified by taxonomic order and an additional analysis was conducted among the various Primate genera. Orders were excluded from the analysis if the total sample size was less than 10 or if no MAYV-positive samples were reported within that order. Pooled seroprevalence was first calculated based on all available data, regardless of test method. This included the samples that tested MAYV-positive based on HI alone (when no confirmatory assay was performed) as well as the samples that were confirmed positive by an NT. Only monotypic reactions to MAYV were included in the meta-analysis in the absence of confirmatory NT. A sensitivity analysis was then conducted using only the MAYV-positive samples that were confirmed using NT. Positive samples that were based on HI alone (without confirmatory NT) were excluded from this analysis, although all MAYV-negative samples were retained. This sensitivity analysis was conducted to account for the low specificity of HI compared to NT [32] and provided a more conservative estimate of seroprevalence.
Due to the substantial differences across studies including sample size, study design, species sampling methods, and geographical location, a random effects model was used for analysis [33, 34]. The Freeman-Tukey double-arcsine transformation was implemented to calculate a proportion, based on the recommendation of Barendregt et al. [35]. A sensitivity analysis was conducted using a generalized linear mixed model (GLMM) with a logit transformation, due to the potential for misleading results with the double-arccosine transformation [36, 37]. Measures of variance ( τ2), heterogeneity (I2), and statistical significance are presented for each random effects model. An additional sensitivity analysis was conducted using a fixed effects model. Results of sensitivity analyses are presented in the Supplementary materials.
The I2 statistic measures inconsistency across study results and is calculated as I2 = 100% x (Q - df) / Q [38]. The I2 statistic ranges between 0% and 100%, where a value of 0% represents no heterogeneity and larger values represent increased heterogeneity. Animal seroprevalence estimates with 95% confidence intervals (CIs) weighted by sample size are presented as forest plots. All analyses were conducted using the ‘meta’ package in R statistical software version 4.0.2 (R Project for Statistical Computing, Vienna, Austria) [39, 40].
Estimation of bias
An assessment of publication bias was carried out for meta-analyses that included five studies or more. Bias was assessed using funnel plots and tests for funnel plot asymmetry based on methods proposed by Egger [41]. If the Egger’s test revealed bias, the Trim and Fill technique was used to estimate the effect of missing studies on the outcomes of the meta-analysis [42].
Georeferencing of MAYV occurrence
All available location information from each confirmed MAYV infection (animal and mosquito) was extracted from each article and georeferenced based on methods that have been described previously [43, 44]. Each occurrence of MAYV was designated as either a point or polygon location according to the spatial resolution provided in the study. When specific latitude and longitude coordinates were provided, they were verified in GoogleMaps and designated as a point location. If a neighborhood, town, village, or small city was explicitly mentioned in the article and fell within a 5×5 km grid cell, it was designated as a point location and its centroid coordinates were recorded. For studies that report a less precise spatial resolution such as states or counties, first level (ADM1) or second level (ADM2) administrative divisions were recorded as polygons. If the size of a specific named location was greater than a 5×5 km grid cell the occurrence was assigned to a custom polygon created in ArcGIS that encompassed the extent of that location. If place names were duplicated (i.e., the ADM1 and ADM2 units had the same name), the coarsest spatial resolution was used. Country shapefiles were accessed through the geoBoundaries Global Administrative Database [45].
Results
General Findings
We identified a total of 57 research items that met our eligibility criteria out of 1523 research items screened, including 46 research articles, seven dissertations, two GenBank entries, one laboratory report, and one abstract (see Table 1 for a full list of eligible items and citations). Thirty-nine (68%) of the included items assessed MAYV infection in non-human vertebrates while 29 (51%) items assessed MAYV infection in arthropods. Of the 57 eligible items, 24 (42%) were included in the vertebrate seroprevalence meta-analysis, and the remaining items were only included in the qualitative analysis. A flow chart describing the article search and selection process is presented in Fig 1. Five articles were identified that met the inclusion criteria but were deemed to be reporting the same data as other included articles. These include de Thoisy et al., (2001) [46] and Talarmin et al., (1998) [12] (both reporting the same data as de Thoisy et al., (2003) [21]), Aitken et al., (1960) [47] (reporting the same data as Aitken et al., (1969) [48]), Batista et al., 2013 [49] (reporting the same data as Paulo et al., (2015) [50]), and Woodall (1967) [51] (reporting the same data as Taylor, (1967) [52]). These articles were excluded from this systematic review.

Studies were conducted in the following countries: Brazil (n=34), Panama (n=5), Colombia (n=4), Peru (n=3), Trinidad and Tobago (n=2), French Guiana (n=1), Venezuela (n=1), Costa Rica (n=1), and the United States of America (n=1). Several studies reported data from multiple countries including Argentina/Paraguay (n=1), Panama/Colombia (n=1), and Brazil/Trinidad and Tobago (n=1). The majority of studies were conducted after the year 2000 (n=33), although some studies were conducted between 1950-1969 (n=9), 1970-1989 (n=8), or 1990-1999 (n=4). Quality scores for all included studies are reported in Table 2.
MAYV in wild-caught non-human vertebrate animals
Thirty-nine (68%) studies in our systematic review assessed MAYV infection in wild-caught non-human vertebrate animals (including birds, mammals, and reptiles). Seventeen (44%) of these studies identified at least one non-human vertebrate that was positive for MAYV infection. Of the 27 taxonomic orders studied, 12 (44.4%) had evidence of MAYV infection: Artiodactyla (even-toed ungulates), Caprimulgiformes (nightbirds), Carnivora, Charadriiformes (shorebirds), Cingulata (armadillos), Columbiformes (pigeons and doves), Didelphimorphia (opossums), Passeriformes (passerine birds), Pilosa (sloths and anteaters), Primate, Rodentia, and Squamata (scaled reptiles). The greatest number of MAYV-positive animal species were found in the order Charadriiformes (n=16 positive species) and the order Primate (n=15 positive species). (See S2 Table for complete mammal data and S3 Table for complete avian data).
Table 3 reports NHP species that were detected with MAYV antibodies. Only studies with positive results are shown on Table 3; other negative studies are listed in the S2 Table. High MAYV seroprevalence was confirmed by NT among Alouatta seniculus monkeys in individual studies in French Guiana [21] (n=51/98) and among Callithrix argentata monkeys in Brazil [19] (n=32/119). In addition, 29 Cebus libidinosus monkeys from wildlife screening centers were detected with MAYV antibodies according to HI, although only six were reported as monotypic reactions [78]. Diagnosis in these monkeys was not confirmed by NT. An additional Cebus libidinosus monkey presented a heterotypic reaction to MAYV (titer of 1:20) and four additional viruses according to HI (including a titer of 1:640 for Oropouche virus) [73]. However, based on the study’s protocol, confirmatory NT was only performed for viruses with titers > 1:40.
Among the 12 additional NHP species with evidence of past MAYV infection, nine were confirmed by NT and three by HI alone. In addition, MAYV positivity was reported in the following NHP genera, although animals were not reported to species: Aotus (n=1/4), Alouatta (n=7/11), Cebus (n=4/13), Sapajus (n=3/43), and Saimiri (n=1/1). The authors reporting MAYV positivity in the Aotus, Alouatta, Cebus, and Saimiri genera noted that these results should be interpreted with caution due to potential for cross-reactivity with Una virus (UNAV) [11]. In one study conducted in Brazil, two of 11 Chikungunya virus (CHIKV)-positive serum samples (in the species Sapajus xanthosternos and Ateles marginatus) neutralized MAYV with titers of 1:40 in plaque reduction NTs [83]. These two samples were considered MAYV-positive and included in our meta-analysis. One additional study [67] detected neutralizing antibodies against both UNAV and MAYV in 21 Alouatta caraya monkeys. However, all 21 monkeys were diagnosed with UNAV based on a 4-fold titer difference between the two viruses. Therefore, we considered these monkeys MAYV-negative and did not include them in our meta-analysis. Finally, in 1963 the Belem Virus laboratory reported MAYV infection in seven NHPs based on HI tests alone [52]. These monkeys were described as Tamarin, Pithecia, and Cebus although no further information was provided regarding sampling method, testing protocol, or primate species.
MAYV antibodies were also detected in 21 bird species from the order Charadriiformes (n=16) and Passeriformes (n=5). All MAYV-positive birds were found in Brazil, with the exception of one MAYV isolate from a migrating bird captured in Louisiana USA [60]. A high MAYV-seroprevalence (n=34/122) was reported by the Belem Laboratory in 1963 among Columbigallina birds, although no additional information was provided regarding sampling method or bird species. MAYV antibodies were also detected in seven avian families that were not identified to genus or species. Only one study that detected MAYV antibodies in birds performed confirmatory NT [19]. All other diagnoses (with the exception of the virus isolation) were made by HI tests alone. See Table 4 for additional information regarding avian species that were infected with MAYV.
Additional wild-caught mammals with evidence of MAYV infection are presented in Table 5. Six rodent species as well as unidentified rodents in the Echimys and Proechimys genera were detected with MAYV antibodies in French Guiana [21], Peru [88], and Panama [20]. In addition, four species in the order Didelphimorphia, three species in the order Pilosa, and one species each in the orders Carnivora, Artiodactyla, and Cingulata were detected with MAYV antibodies in French Guiana [21] and Peru [88]. Additional positive samples were detected in the orders Rodentia, Didelphimorphia, and Pilosa although the species were not identified.
Successful isolation of MAYV was reported from the following viremic animals: a silvery marmoset (Callithrix argentata) captured during a MAYV outbreak in Belterra, Brazil [19] and a migrating orchard oriole (Icterus spurius) captured in Louisiana [60]. In addition, the Belem Virus Laboratory reported MAYV isolation from two lizard species in 1963 [52] (Tropidurus torquatus and Ameiva ameiva) although no further information was provided regarding study methods or procedures.
The geographic distribution of animals (wild-caught, domestic, and sentinel) infected with MAYV is presented in Fig 2. The infected animals were identified in six countries overall, including Brazil, Peru, French Guiana, Colombia, Venezuela, and Panama, although the majority of infected animals were found in Brazil. Overall, 12 locations were geo-referenced as points, four locations as ADM1 polygons, 15 locations as ADM2 polygons, and two locations as custom polygons.
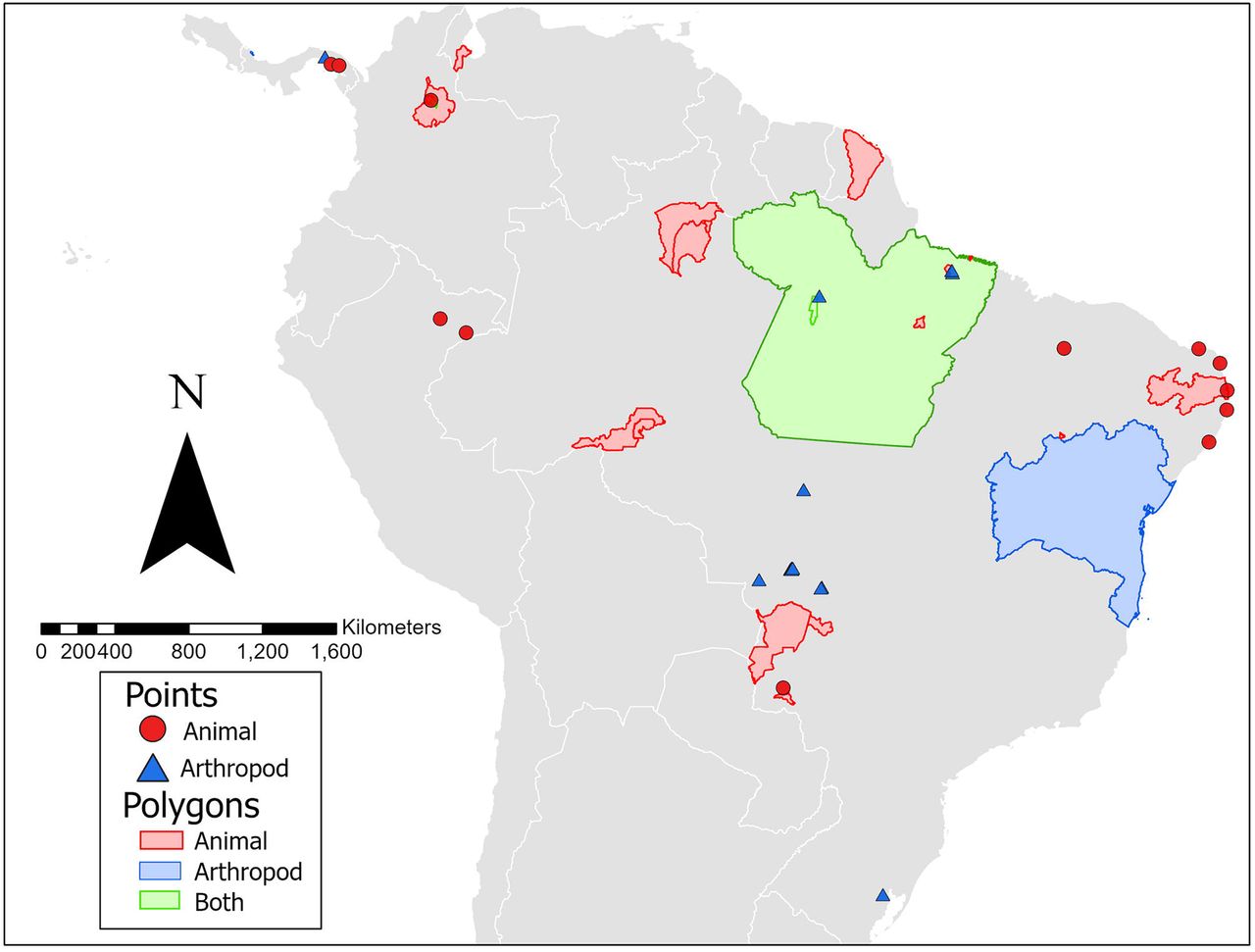
The finest spatial scale is presented where possible. One MAYV isolate detected in a migrating bird in Louisiana is not included in the map.
MAYV in domestic or sentinel animals
Nine studies analyzed MAYV seroprevalence in domestic animals (equids, sheep, poultry, dogs, pigs, cattle, and buffaloes), and five studies analyzed MAYV seroprevalence in sentinel animals (monkeys, mice, and hamsters). Domestic and sentinel animals with evidence of MAYV positivity are reported in Table 6 and complete results are reported in the S4 Table. In domestic animals, evidence of MAYV infection was detected in equids, cattle/buffalo, and dogs. Six studies assessed MAYV seroprevalence in Brazilian equids [54, 57, 63, 74, 85, 86], and antibodies against MAYV were detected in four of these studies. Notably, Gomes et al. [74] reported MAYV neutralizing antibodies in 48 equids out of 213 (23%) based on ELISA. However, only 16 of the 48 equids were considered positive based on the study’s diagnostic criterion of 4-fold greater plaque reduction NT90 titer than that of the other viruses under study. In addition, Casseb et al. [63] detected MAYV antibodies in 40 horses using HI, although only four of the 40 reactions were monotypic, and confirmatory NTs were not performed. Additional domestic animals with evidence of MAYV infection included cattle/buffalo (n=14/1103 positive reactions by HI; 5/14 monotypic reactions [62]) and dogs (n=2/7 positive reactions by HI [53]). In addition, neutralizing antibodies (plaque reduction NT90 titer >10) against MAYV were detected in three sheep in Brazil [86]. However, these animals did not meet the original study’s diagnostic criterion for MAYV diagnosis based on 4-fold greater plaque reduction NT90 titer than that of the other viruses under study. Evidence of MAYV infection was also detected by HI in two sentinel monkeys placed in the tree canopy in Panama [98], and one MAYV isolate was obtained from a sentinel hamster in Venezuela [82].
Pooled prevalence of MAYV in non-human vertebrate animals
Twenty-four studies overall were included in the pooled prevalence meta-analysis. Eight studies were excluded because they did not clearly state how many animals were tested for MAYV within each order [55, 66, 72, 84, 89, 93] or did not present serologic results [60, 70]. Another study was excluded because authors reported the number of “Group A” positive serum samples, but did not specify individual viruses [98]. Studies were also excluded if they only reported sequence data or only included sentinel animals [82, 91, 95, 100]. Finally, a study that sampled bats exclusively was excluded because no MAYV-positive samples were reported in the order Chiroptera [92].
Eleven orders of nonhuman vertebrate animals (including domestic equids) were included in the meta-analysis. Orders were excluded from the analysis due to insufficient sample size (N<10) or if no MAYV-positive samples were reported. These include the orders Apodiformes (MAYV prevalence: 0/3), Caprimulgiformes (MAYV prevalence: 1/6), Chiroptera (MAYV prevalence: 0/1546), Crocodilia (MAYV prevalence: 0/87), Cuculiformes (MAYV prevalence: 0/5), Galliformes (MAYV prevalence: 0/1), Gruiformes (MAYV prevalence: 0/2), Psittaciformes (MAYV prevalence: 0/3), Tinamiformes (MAYV prevalence: 0/2), Pelecaniformes (MAYV prevalence: 0/2), and Podicipediformes (MAYV prevalence: 0/2).
The primate order appeared in 14 studies that were included in the meta-analysis. When all positive samples were included, the pooled MAYV seroprevalence among primates was 13.1% (95% CI: 4.3-25.1%) according to the random effects model, with statistically significant heterogeneity across studies (I2 = 95%, p < 0.01). After excluding positive samples that were not confirmed by NT, the pooled MAYV seroprevalence among primates decreased to 4.9 (95% CI: 0.0-15.2; I2 = 96%; p < 0.01) according to the random effects model. When the analyses were repeated using the GLMM with logit transformation, seroprevalence estimates for primates decreased to 8.7% (95% CI: 3.1-22.0%) overall and to 0.7% (95% CI: 0.0-9.1%) when only NT-positive samples were included. Additional meta-analysis results for the various primate genera are presented in S6 and S7 Tables. The seroprevalence for the most frequently sampled primate genera was 32.2% (95% CI: 0.0-79.2%) for the Alouatta genus, 17.8% (95% CI: 8.6-28.5%) for the Callithrix genus, and 3.7% (95% CI: 0.0-11.1%) for the Cebus/Sapajus genus.
Meta-analysis results for additional non-human vertebrate orders are presented in Table 7 and forest plots for mammal orders and avian orders are presented in Figs 3 and 4, respectively. When all positive samples were included in the analysis, the highest seroprevalence was observed in the orders Charadriiformes (prevalence: 7.1%; 95% CI: 2.2-13.8%) and Cingulata (prevalence: 3.0%; 95% CI: 0.0-24.5%). When the analysis was repeated using GLMM with logit transformation, the seroprevalence increased to 10.0% (95% CI: 2.7-30.8%) for the order Cingulata and 9.2% (95% CI: 4.4-18.2%) for the order Charadriiformes. All results of the sensitivity analysis using GLMM with logit transformation are reported in the S5 Table. An additional sensitivity analysis using fixed effects models is presented in the S8 and S9 Tables.

Estimates are based on random effects model with Freeman-Tukey double arcsine transformation. All samples that tested MAYV-positive are included, regardless of test method.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Estimates are based on random effects model with Freeman-Tukey double arcsine transformation. All samples that tested MAYV-positive are included, regardless of test method.
MAYV in wild-caught arthropods
Twenty-eight of the studies in our systematic review analyzed MAYV infection in wild-caught arthropods. Seventeen (61%) of the 28 studies reported at least one arthropod that was positive for MAYV infection. Of the mosquito genera studied, seven were found to be infected with MAYV: Aedes, Culex, Haemagogus, Psorophora, Sabethes, Wyeomyia, and Mansonia. For detailed information on all infected mosquito species, see Table 8. The majority of infected vectors were identified using viral isolation techniques, although three studies reported MAYV positivity using RT-PCR alone. In addition, one study reported isolation of MAYV from an Ixodes tick [91] while another study reported isolation from a Gigantolaelaps mite [52]. Complete results, including studies that did not detect MAYV in arthropods, are reported in the S10 Table.
The geographic distribution of vectors infected with MAYV is presented in Fig 2. MAYV-positive arthropods were identified in four countries overall, including Brazil, Colombia, Panama, and Trinidad. Overall, 15 locations were geo-referenced as points, two locations as ADM1 polygons, two locations as ADM2 polygons, two locations as ADM3 polygons, and two as custom polygons.
Analysis of publication bias
Publication bias was assessed among six animal orders (including domestic equids) and two primate genera. The results of Egger’s test did not reveal evidence of publication bias for the included studies. Therefore, the Trim fill technique was not carried out. Funnel plots are presented in S1 and S2 Figures, and results of Egger’s test are reported in the S11 Table.
Discussion
To our knowledge, this study is the first attempt to systematically review the existing evidence of non-human animal reservoirs and arthropod vectors of MAYV, and the first study to quantitatively analyze the pooled seroprevalence of potential reservoirs. We identified 57 studies that assessed MAYV infection in non-human vertebrate animals and arthropods. Overall, the studies found evidence of MAYV infection in 12 wild-caught animal orders and seven arthropod genera across seven Latin American countries and the USA.
The majority of animal species that were found to be infected with MAYV belonged to the orders Primate and Charadriiformes (shorebirds). Several MAYV-positive species were also detected in the orders Rodentia, Didelphimorphia, and Pilosa. Overall, the highest MAYV pooled prevalence occurred in the Primate order. This finding points to the potential role of NHPs as an important reservoir in the MAYV transmission cycle.
The role of NHPs in sylvatic transmission cycles of arboviruses has been demonstrated with varying degrees of evidence [101]. Several arboviruses have been successfully isolated from wild NHPs, including dengue [102], CHIKV [103], and Zika [104] viruses. While isolation of a virus from NHPs is important for establishing the existence of a sylvatic cycle, it is difficult to achieve due to the short duration of viremia [101]. In our review, we identified only one study that successfully isolated MAYV from a NHP [19]. In the absence of viral detection, antibody seroprevalence has been used as evidence of the role of NHPs in sylvatic transmission cycles [105, 106]. Therefore, the high seroprevalence of MAYV among NHPs, including 52% seropositivity among A. seniculus monkeys in a 1994-95 survey in French Guiana [21], points to the potential importance of NHPs as MAYV reservoirs. Furthermore, Hoch et al. [19] reported substantial viremia in C. argentata marmosets that were experimentally infected with MAYV and noted that viremia titer was likely sufficient to infect vectors. Due to the high MAYV seroprevalence among marmosets during the Belterra outbreak, the isolation of MAYV from a single C. argentata marmoset, and the results of experimental infection studies, the authors concluded that marmosets were likely the amplifying hosts of MAYV.
The importance of birds in the MAYV transmission cycle was hypothesized following viral isolation from a migrating oriole (Icterus spurius) in Louisiana [60]. Avian species have been implicated as definitive or potential reservoirs of several Alphaviruses, including Sindbis virus [107], Ross River virus [108], and Eastern/Western equine encephalitis virus [109]. However, their role in MAYV transmission remains poorly understood. Our systematic review identified seven studies that found MAYV positivity in birds in the orders Passeriformes, Caprimulgiformes, Columbiformes, and Charadriiformes with relatively high seroprevalence reported in several bird species in the latter two orders [52, 53]. While some have theorized that MAYV has been introduced into certain areas by migratory birds [59], this hypothesis requires further study in order to elucidate the role of birds in MAYV transmission.
Although evidence of MAYV infection was detected in several vertebrate species, identifying the primary non-human animal reservoirs remains a difficult task. The precise definition of a disease “reservoir” has been a source of disagreement [17, 110]. One definition proposed by Haydon et al., (2002) defined a reservoir as “one or more epidemiologically connected populations or environments in which the pathogen can be permanently maintained and from which infection is transmitted to the defined target population” [17]. In addition, in 2005 Kuno and Chang outlined three basic criteria for the identification of reservoirs including isolation of the virus from the suspected reservoir population, high antibody prevalence in field-caught animals, and evidence of viremia in laboratory settings, although they posited that definitive identification of a reservoir requires evidence of long-term infection [111]. The role of various non-human vertebrates in the MAYV transmission cycle should be explored further in longitudinal seroprevalence surveys and experimental transmission studies in laboratory settings.
The sylvatic Hg. janthinomys mosquito has long been considered as the primary vector of MAYV. This is in part based on the isolation of MAYV from several pools of Hg. janthinomys mosquitoes in the context of a major MAYV outbreak in Belterra, Brazil in 1978 [19]. Our systematic review also identified several additional mosquito species including Ae. aegypti and Cx. quinqefasciatus with evidence of MAYV infection. A caveat, however, is that the isolation of a virus or detection of viral RNA through PCR is not sufficient to establish that arthropod as a biological vector [112], i.e. involved in the biological transmission of pathogens [111]. The World Health Organization (WHO) established three criteria to define a confirmed vector: (1) viral isolation in the absence of vertebrate blood; (2) biological transmission of the virus in experimental conditions; and (3) presence of certain temporal, geographic and other epidemiological or ecological parameters that allow transmission to occur [112]. Thus, certain arthropods that are capable of ingesting and transmitting a virus may not be established as confirmed vectors if the other parameters are not in place.
Experimental transmission studies support the role of Ae. aegypti as a possible MAYV vector with high MAYV infection rates and transmission potential [22-24, 113]. For example, Long et al., revealed Ae. aegypti to be a capable MAYV vector with a relatively short extrinsic incubation period [22]. Furthermore, MAYV titers in the saliva of Ae. aegypti were similar to other Alphavirus-vector systems including EEEV in Culiseta melanura and VEEV in Ae. albopictus and Ae. taeniorhynchus. In contrast, Cx. quinquefasciatus mosquitoes exhibited low MAYV infection rates and inability to transmit MAYV in laboratory settings [113]. It is also important to note that the competence of a given vector species to transmit MAYV may be impacted by the MAYV genotype that is present in a given area. In laboratory conditions, genotype L infection rates were significantly higher than genotype D infection rates among Ae. aegypti mosquitoes [113].
The spillover of MAYV into urban populations has been a source of concern for Latin American health authorities [114]. The implication is that anthropophilic, urban-dwelling mosquitoes like Ae. aegypti as effective vectors of MAYV would increase the potential for urban MAYV outbreaks [115]. Concerns of urban MAYV transmission were amplified after antibodies to MAYV were discovered in 33 of 631 sera (5.2%) in the city of Manaus, Brazil in 2007-08 [25] although it is unclear if humans can serve as amplification hosts. For example, Long et al. noted that the short duration of MAYV viremia and the relatively low viremic titers in humans reduces the probability of urban spread [22]. Our systematic review identified two recent studies conducted in the city of Cuiaba in which MAYV was isolated from pools of wild-caught Ae. aegypti mosquitoes [79, 96]. One of these studies also reported vertical transmission of MAYV [79]. This represents another mechanism that may lead to maintenance of the virus in urban mosquito populations. Although Ae. aegypti mosquitoes have not been conclusively implicated as MAYV vectors, the isolation of MAYV from wild-caught Ae. aegypti mosquitoes combined with the evidence of vector competence in laboratory settings [22-24, 113] suggests that MAYV could spill over into an urban cycle. This hypothesis requires further study to explore natural MAYV infection in city-dwelling mosquitoes and additional controlled vector competence studies.
Our systematic review revealed substantial heterogeneity across included studies, even within animal orders. Heterogeneity may complicate the interpretation of pooled seroprevalence estimates [38]. An additional limitation involves the validity of serological assays used to detect MAYV infection in animals. While plaque reduction NT is considered the “gold standard test” for detecting neutralizing antibodies to MAYV, some of the studies in the review instead relied on the less-specific HI test for antibody detection [101]. Furthermore, antibodies to other alphaviruses in the Semliki Forest serocomplex (e.g., CHIKV) may cross-react in serological tests [116]. Therefore, interpretation of seroprevalence estimates should be done with caution especially in the absence of confirmatory NT. Finally, unpublished data and articles with low quality scores were included in this review due to the paucity of eligible studies. Therefore, readers should consider the heterogeneity of study quality when interpreting the results of pooled seroprevalence estimates.
Conclusions
MAYV is an emerging arbovirus that poses a major threat to human populations in Latin America. In order for public health authorities to effectively design MAYV surveillance and control programs, an understanding of the disease ecology is essential. This systematic review adds to existing knowledge regarding the potential animal reservoirs and arthropod vectors that are involved in the MAYV transmission cycle. These baseline data and maps of MAYV occurrence can direct risk emergence modeling and prediction efforts. Future studies involving experimental infection of primates and other non-human vertebrates are necessary to determine the animal species that may serve as amplifying hosts. Furthermore, additional experimental transmission studies may provide critical information regarding the potential for Ae. aegypti to facilitate urban spread of MAYV.
FUNDING STATEMENT
This work was in part conducted by the Infectious Disease Clinical Research Program (IDCRP), a Department of Defense (DoD) program executed by the Uniformed Services University of the Health Sciences (USU) through a cooperative agreement with The Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc. (HJF). This project has been supported with federal funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health (NIH), under Inter Agency Agreement Y1-Al-5072 and from the Defense Health Program, U.S. Department of Defense, under award HU0001190002.
A.A.’s participation in this work was supported by funding from Armed Forces Health Surveillance Branch - Global Emerging Infections Surveillance (GEIS) Project #P0044_20_NS and NASA Applied Sciences Program – Health and Air Quality, Grant #17-HAQ17-0065.
DISCLAIMER
The contents of this publication are the sole responsibility of the author(s) and do not necessarily reflect the views, opinions or policies of Uniformed Services University of the Health Sciences (USUHS), the Department of Defense (DoD), the Departments of the Army, Navy, or Air Force. Mention of trade names, commercial products, or organizations does not imply endorsement by the U.S. Government.
CONFLICT OF INTEREST
The authors declare no conflicts of interest.
Supplementary Materials
S1 Table. PRISMA Checklist
S2 Table. MAYV positivity by taxa of wild mammals in included studies S3 Table. MAYV positivity by taxa of wild birds in included studies
S4 Table. MAYV positivity in domestic or sentinel animals studied
S5 Table. Pooled Prevalence Table (Random effects using GLMM with logit transformation)
S6 Table. Primate Genera Pooled Prevalence Table (Random effects with Freeman-Tukey double arcsine transformation)
S7 Table. Primate Genera Pooled Prevalence Table (Random effects using GLMM with logit transformation)
S8 Table. Pooled Prevalence Table (Fixed effects with Freeman-Tukey double arcsine transformation)
S9 Table. Pooled Prevalence Table (Fixed effects using GLMM with logit transformation) S1 Fig. Funnel plots for estimates of MAYV seroprevalence in non-human animal reservoirs
S2 Fig: Funnel plots for estimates of MAYV seroprevalence in non-human primate genera
S10 Table. Complete arthropod results by genus
S11 Table. Egger’s test for publication bias
Acknowledgments
We would like to thank Dr. Mauro Ramos for his assistance with reviewing Portuguese language articles. ELE is a Scientific researcher of the Consejo de Investigaciones Científicas y Tecnológicas (CONICET) from Argentina.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.
- 57.↵
- 58.
- 59.↵
- 60.↵
- 61.
- 62.↵
- 63.↵
- 64.
- 65.
- 66.↵
- 67.↵
- 68.
- 69.
- 70.↵
- 71.
- 72.↵
- 73.↵
- 74.↵
- 75.
- 76.
- 77.
- 78.↵
- 79.↵
- 80.
- 81.
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.
- 88.↵
- 89.↵
- 90.
- 91.↵
- 92.↵
- 93.↵
- 94.
- 95.↵
- 96.↵
- 97.
- 98.↵
- 99.
- 100.↵
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.↵
- 109.↵
- 110.↵
- 111.↵
- 112.↵
- 113.↵
- 114.↵
- 115.↵
- 116.↵




