

Abstract
The neurodevelopmental phenotype in Down Syndrome (DS), or Trisomy 21, is variable including a wide spectrum of cognitive impairment and a high risk of early-onset Alzheimer’s disease (AD). A key metabolite of interest within the brain in DS is Myo-inositol (Myo-ins). The NA+/Myo-ins co-transporter, is located on human chromosome 21 and is overexpressed in DS. In adults with DS, elevated brain Myo-ins has previously been associated with cognitive impairment and proposed as a risk marker for progression to AD. However, it is unknown if brain Myo-ins is increased earlier in development.
The aim of this study was to assess Myo-ins and key brain metabolites [N-acetylaspartate (NAA), Choline (Cho) and Creatine(Cr)] in the developing brain in DS and aged-matched controls. To achieve this we used mass spectrometry in early (10-20 weeks post conception) ex vivo fetal brain tissue samples from DS (n=14) and control (n=30) cases; and in vivo magnetic resonance spectroscopy (MRS) in neonates with DS (n=18) and aged matched controls (n= 25) scanned just after birth (36-45 weeks postmenstrual age.
We observed elevated Myo-ins in the ex vivo fetal cortical brain tissue in DS compared with controls. Relative to reference metabolites Cho and Cr, we also detected elevated ratios of Myo-ins and NAA in vivo in the basal ganglia and thalami, in neonates with DS, when compared to age-matched typically developing controls. Thus, a higher level of brain Myo-ins was evident as early as 10 weeks post conception and was measurable in vivo from 36 weeks post-menstrual age. Future work will determine if this early difference in metabolites is linked to cognitive outcomes in childhood or has utility as a potential treatment biomarker for early intervention.
Introduction
Down Syndrome (DS) is a complex genetic condition, resulting from the triplication of human chromosome 21 (HSA21; (Trisomy 21), and is associated with a wide spectrum of neurodevelopmental outcomes (Karmiloff-Smith et al., 2016; Baburamani et al., 2019). Individuals typically present with cognitive deficits, and variable impairments in speech, language and motor functions, coupled with a high risk of early-onset dementia and Alzheimer’s disease (AD) (Rachidi and Lopes, 2007; Wiseman et al., 2015; Karmiloff-Smith et al., 2016). There is limited understanding of brain development in DS, but early alterations may contribute to the variations in lifelong neurodevelopmental outcomes and ultimately to the risk for early-onset AD. Recent in vivo studies have demonstrated altered structural brain development in DS from as early as 20 weeks gestation, but it is not known whether these are associated with abnormal metabolic profiles (Patkee et al., 2020). Metabolic derangements, specifically with an elevation of myo-inositol (myo-ins), have been described across development in mouse models of DS, as well as in the child and adult human brain with DS, but there have been no studies assessing brain metabolism at much younger ages.
Myo-ins is of particular interest as the NA+/Myo-inositol co-transporter (SLC5A3) is located on HSA21 and is overexpressed in DS (Berry, Wang and Dreha, 1999). Magnetic resonance spectroscopy (MRS) studies in the adult DS population have reported significantly elevated ratios of myo-ins in DS brains, with and without the neuropathological hallmarks of AD, as compared with healthy adult controls (Lin et al., 2016). Higher levels of Myo-ins have been found to correlate with poor cognitive performance, suggesting that this metabolite might have utility for monitoring response to treatment and/or constitute a treatment target (Lin et al., 2016). Consistent with the latter, studies in mouse models of DS have shown that higher levels of Myo-ins could be reduced by treatment with lithium, which also rescued both synaptic plasticity and memory deficits (Huang et al., 1999; Contestabile et al., 2013). Whilst such studies have made valuable first steps to revealing a link between the metabolite profile of DS and functional outcomes, it is unknown when this alteration in Myo-ins (and/or other metabolites supporting brain function) emerges.
Current therapeutic approaches to improve cognition in DS, target children and adults; and these have had limited success. For example, a clinical trial of lithium in adults with DS did not report efficacy (Murphy, 2011). Arguably however, intervention may be needed at an earlier timepoint. Identification of neural differences much earlier in the life course in DS, could potentially offer opportunities for more effective intervention. We and others have recently demonstrated that fetuses and neonates with DS have reduced whole brain, cortical and cerebellar volumes from as early as 20 weeks gestational age (GA), using in vivo volumetric MRI (Tarui et al., 2019; Patkee et al., 2020). Whether such anatomical differences are accompanied by early differences in the metabolite levels, that reflect neuronal and glial function, and alter neurodevelopmental outcome, remains unknown.
The aim of this study was to determine whether the brain metabolite profile and Myo-ins, specifically, is already altered in DS in during early development in utero. We used in vivo MRS to assess the metabolic profile of brain tissue (basal ganglia and thalami) in neonates with DS compared to typically developing controls. To complement this, we used data from a second study and analysed the metabolite profile in post-mortem fetal cortical brain tissue using Mass Spectrometry. In addition to Myo-ins, other metabolites of interest we measured were N-acetylaspartate (NAA), Choline (Cho) and Creatine (Cr). We hypothesised that there would be significant alterations in metabolic ratios between the DS and control groups, with increases in Myo-ins levels specifically, in the brain of neonates with DS and secondly, increases in absolute Myo-ins at earlier gestations in brain tissue from foetuses with DS compared with controls
Methods
Magnetic Resonance Spectroscopy Study
Ethical approval for this study was obtained from the West London and GTAC Research Ethics Committee (REC) for DS participants (07/H0707/105); and from the Dulwich NREC for the control population (12/LO/2017). Informed consent was obtained from the legal guardians of all infants, prior to imaging at St. Thomas’ Hospital, London, UK. All methods were carried out in accordance with the relevant guidelines and regulations.
Participants
Participants who had previously had a fetal scan (Patkee et al., 2020) and consented during their initial scan to be contacted post-delivery, were invited for a neonatal scan up to 46 weeks post-menstrual age (PMA). Neonates with confirmed DS were also recruited from the neonatal unit or postnatal wards at St Thomas’ Hospital. Participants with DS, some of whom had other non-brain congenital abnormalities such as cardiac defects or gastrointestinal malformations, were included in this study. Clinical details for the neonates with DS can be found in Table 1. Control group neonates were recruited from South London and South East of England antenatal centres as part of the Brain Imaging in Babies Study (BIBS) and scanned at the Centre for the Developing Brain (St. Thomas’ Hospital, London, UK). All infants were scanned on the same scanner with identical imaging protocols. Neonates were included as controls if they had a normal brain appearance on the neonatal MRI, with no other congenital or chromosomal abnormalities, and an uneventful delivery, as acute birth-related events could potentially change metabolite composition even in the presence of normal MR imaging. Additionally, all control neonates had no known immediate family members with any neurodevelopmental or behavioural conditions (e.g. autism and ADHD). Previous MRS studies in the paediatric population found statistically significant differences between DS and control groups with 20 participants in each cohort (Śmigielska-Kuzia et al., 2010).
Gestational age (GA), post-menstrual age (PMA), atrioventricular septal defect (AVSD), atrial septal defect (ASD), ventricular septal defect (VSD) and patent ductus arteriosus (PDA)
Neonatal Image Acquisition
MR scanning of the DS and control neonates was performed on a Philips Achieva 3-Tesla system (Best, The Netherlands) at the Centre for the Developing Brain (St. Thomas’ Hospital, London, UK) using a dedicated 32 channel neonatal head coil (Hughes et al., 2017). Infants were placed in supine and secured within the scanning shell and a neonatal positioning device was used to stabilise the head to reduce movement (Hughes et al., 2017). Auditory protection was comprised of earplugs moulded from silicone-based putty placed in the outer ear (President Putty, Coltene/Whaledent Inc., OH, USA), neonatal earmuffs over the ear (MiniMuffs, Natus Medical Inc., CA, USA) and an acoustic hood placed over the scanning shell. Sedation was not administered, and all babies were scanned during natural sleep, following a feed. An experienced neonatologist or neonatal nurse was present during the examination and pulse oximetry, heart rate, and temperature was monitored throughout the scan.
T2-weighted images were acquired in the coronal, sagittal and transverse planes using a multi-slice turbo spin echo sequence. Two stacks of 2D slices were acquired using the scanning parameters: TR = 12 s; TE = 156 ms; slice thickness = 1.6 mm with a slice overlap = 0.8 mm; flip angle = 90 degrees; matrix /field of view= 320 x 320 x 125; and an in-plane resolution: 0.8×0.8 mm. All T2-weighted images were reviewed to assess image quality and those with excessive motion and uninterpretable spectra were excluded. Visual analysis of all neonatal MR images was performed by a specialist perinatal radiologist to exclude additional anomalies and confirm appropriate appearance for gestation. Image datasets showing an overt additional structural malformation or brain injury not considered to be part of the expected DS phenotype were excluded. The data regarding T2-weighted volumetric findings are detailed in Patkee et. al., (2020).
Spectral Acquisition
A PRESS sequence was used to obtain the MR spectra in the neonatal brains. A region of interest 20 x 20 x 20 mm3 was placed centrally over the left basal ganglia and thalami to acquire spectroscopy (Fig 1). Previous studies on early brain development have observed higher individual metabolite levels in the deep grey matter, compared to other regions such as frontal lobe white matter (Kim, Barkovich and Vigneron, 2006; Tomiyasu et al., 2013; Lally et al., 2019). This is additionally beneficial due to the reduced likelihood of contamination by scalp fat. MRS uses a double spin echo sequence (90° – 180° – 180°) to improve signal quantity.
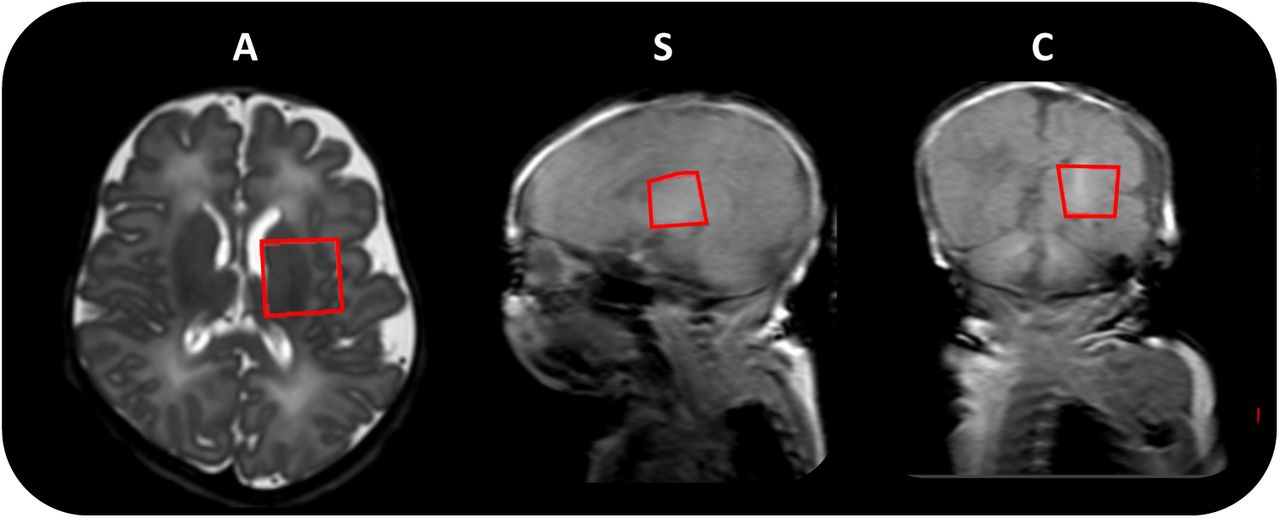
Region of interest size 20 x 20 x 20 mm3 (left basal ganglia and thalami) indicated by the red box, in the axial (A), sagittal (S) and coronal (C) planes respectively.
Each spectrum took approximately 3 minutes to obtain and was acquired in blocks of four averages (producing 32 individual spectra) and saved independently, allowing for the elimination of individual corrupted spectra as a result of excessive motion or contamination of peaks, prior to post-processing the data. Water suppression was performed by selective excitation. This is necessary to avoid metabolite signals being overwhelmed by the large water peak and to ensure that the baseline of the spectrum is not distorted, which can make it difficult to quantify individual smaller peaks. In this study, metabolites of interest were measured at two echo times, TE 55 and 144 ms.
Processing and Analysis of Spectral Data
The software installed on the Philips MRI scanner performs some basic rapid automatic processing for immediate quality assessment of the acquired spectrum. For a complete quantitative analysis of metabolite ratios, spectra were post-processed offline using the jMRUI software package (http://www.jmrui.eu/, version 3.0. The spectral analysis aims to estimate signal intensities or areas under the peaks corresponding to the metabolites. Individual spectra were quality assessed for high SNR by the height of metabolite peaks as compared to background noise. Data with significant motion or those with aberrant water peaks were discarded. The remaining spectra with a residual water peak at the accepted threshold were summed together to produce a final spectrum, after which the water peak at 4.7ppm was removed (Fig 2). In this study we focused on the following metabolites of interest (resonant frequency); N-acetylaspartate (2.0 ppm), Creatine (3.0 ppm), Choline (3.2 ppm) and Myo-inositol (3.5 ppm).

using a PRESS sequence at an echo time of 144ms. Peaks for Myo-inositol (3.5 ppm), Choline (3.2 ppm), Creatine (3.0 ppm) and N-acetylaspartate (2.0 ppm), are labelled, following the removal of the H2O peak.
Quantification of Metabolites
The jMRUI software allows for the time-domain analysis of MR spectra using the AMARES programme. Expected peaks for each of the metabolites were identified on the spectrum based on their frequency (ppm). The top and midpoint of each peak were identified and based on this prior knowledge a curve was fit by the software for each metabolite. Once the curves had been fitted, metabolites ratios were calculated for the two echo times as follows: TE 55 ms; Myo:Cho, Myo:Cr, Cho:Cr and TE 144 ms; NAA:Cho, NAA:Cr, Cho:Cr.
The different peaks in the spectrum represent individual metabolites and are identified by the frequency at which they resonate, represented in parts per million (ppm). In MRS, the total area under a metabolite resonance in an MR spectrum is directly proportional to the concentration of the metabolite. As the MR signal is affected by factors such as relaxation time and the acquisition method, amongst others, it is not possible to establish an absolute concentration without accounting for these confounding elements. At present, metabolites are usually calculated as a relative quantification and therefore are expressed as ratios rather than as absolute concentrations. This is particularly useful when detecting subtle or more sensitive changes and can be more accurate than an absolute quantification; this is true in cases with pathology where one metabolite concentration rises whilst the other declines (Jansen et al., 2006). All neonatal spectra were processed by the same operator.
Mass Spectrometry Study
Fetal Brain Tissue Samples
Human fetal cortex material was provided by the Joint MRC/Wellcome Trust (MR/R006237/1) Human Developmental Biology Resource (HDBR) (www.hdbr.org). HDBR provided fresh fetal cortex tissue that was snap-frozen for DS (n=14) and age-matched controls (n=30) in the fetal age range 10 to 20 post-conception weeks (PCW). Genetic diagnosis confirmed presence of Trisomy 21 (DS). There were no other known abnormalities in either group. HDBR specific ethics approval (18/LO/0822, 18/NE/0290) was sought and tissue was collected following termination of pregnancy, following Human Tissue Act (HTA) regulations. Cortical tissue was then selected, snap frozen and stored at -80 °C. Cases that were used are detailed in Table 2.
Age at birth, in post-conceptional weeks (PCW), sex and other genetic/co-morbidities are listed.
Sample preparation
Human fetal cortex tissue samples were stored at –80°C. The cortical tissue used included a variety of regions, as due to the nature of these fetal samples it is often difficult to identify the exact cortical region tissue originates from across the gestational ages. To ensure this did not affect the results, cortical tissue was randomly assigned for the mass spectrometry analysis in both DS and control groups. Approximately 0.1g of brain tissue from the cortex was collected on dry ice and weighed. 1 ml of HPLC grade Methanol (Sigma, UK) was added and samples were then homogenised for 20 minutes at 30 Hz using TissueLyser II device and centrifuged at 14,000 rpm for 0.5 minute (longer if required). Clear supernatants were then transferred to a new tube on dry ice, samples were then stored at -80 °C until analysis. Analysis of Myo-Inositol and NAA was performed with Gas Chromatography Mass Spectrometry (GCMS) and for Creatine and Choline using Liquid Chromatography Mass Spectrometry (LCMS). All metabolite values for an individual fetal sample were measured from a single tissue sample to eliminate inter-assay variability. All analysis was run by the Mass Spectrometry Core Facility, Kings College London (KCL, UK).
Gas Chromatography Mass Spectrometry (GCMS) for Myo-Inositol and NAA
Samples (10ul; Blank, Calibration standard or test samples) were added to tubes containing internal standard-200 (20 µl) + solution (50µl) (mixture of internal standards (200 ug/ml Myo-inositol internal standard, 50 ug/ml NAA internal standard solution). These were evaporated to nearly dry under N2 at 40±5°C. Pyridine (10 µl) and BSTFA (40 µl) were then added and the samples were vortexed, heated at 80±5°C for 30 minutes, cooled down to room temperature and transferred to injection vials. Calibration ranges were 50-2000 µg/ml for Myo-Inositol (Cat# M01914, Fluorochem Limited, UK; Internal Standard Myo-inositol-D6 Cat# 1665997, Toronto Research Chemicals, Canada) and 1-100 µg/ml for NAA (Cat# 00920, SLS, UK). Samples were analysed on an Agilent GCMSD system. GC-MS parameters are detailed in Table 3.
Liquid Chromatography Mass Spectrometry (LCMS) for Creatine and Choline
Samples (100 µl; Blank, Calibration or test samples) were added to individual injection vials containing Internal Standard-2u (50 µl; 2µg/mL Acetylcholine-D4, Cat#D-1555, CDN Isotopes, Canada) and DS1 (900 µl, contains 0.1% Formic Acid in 50 % HPLC grade Acetonitrile), vortexed, and injected into the LCMS system. DS1 was run between high and low concentration samples and between sets of the test sample. Calibration ranges were 10 – 200 µg/ml for Creatine (Cat# BS-9561E, BioServ UK Limited) and 0.10-35.0 µg/ml for Choline (Cat# 67-48-1, Sigma Aldrich, UK). Samples were analysed on a Thermo Accela Pump and CTC Auto sampler coupled to a Thermo TSQ Quantum Access. LC Solvent A (0.1% Formic Acid in water); LC Solvent B (0.1% Formic Acid in Acetonitrile). LCMS parameters detailed in Table 3.
Statistical Analysis
Statistical analysis was performed using MATLAB (Release: 2017a, The MathWorks, Inc., Natick, MA, USA), and visualised in python 3.7 (www.python.org). Linear mixed-effects models (LME) were used to compare results between the two groups (DS and controls) in both the mass spectrometry and MRS studies, controlling for the covariate PMA at scan as a fixed effect. Cohen’s d values were calculated to quantify effect sizes and the magnitude of interactions (Cohen, 1988; Rosenthal and Rosnow, 2008). Effect sizes were interpreted as small (Cohen’s d value ≤ 0.4), medium (0.5-0.7) and large (≥ 0.80) (Cohen, 1988). A post-hoc False Discovery Rate (FDR) test was performed for multiple comparisons, using the Benjamini-Hochberg (1995) procedure, controlling the alpha error to 5%.
Results
Neonatal Magnetic Resonance Spectroscopy
A total of 21 neonates with DS were scanned (Patkee et al., 2020), 18 had usable and interpretable spectral acquisitions. For comparison, 25 control neonates were included as part of this study. Of these, 21 neonates had interpretable spectral acquisitions (Table 4).
Gestational age (GA) and post-menstrual age (PMA).
The Myo:Cho and Myo:Cr ratios in the control cohort appeared to decrease with advancing PMA at scan. There was marked variability in Myo-ins ratios across PMA in the DS population, with no obvious decrease with PMA at scan. Myo:Cho and Myo:Cr ratios were elevated in DS neonates compared to controls, corrected for PMA at scan, with group differences of large effect sizes (p < 0.05, Table 5, Fig 3A, 3B). NAA:Cho and NAA:Cr ratios were found to increase with advancing PMA in both the DS and control groups. There were increased ratios of NAA:Cho and NAA:Cr in the DS cohort compared with controls (p < 0.05, Table 5, Fig 3C, 3D). Cho:Cr ratios decreased with increasing PMA in both cohorts. The significance of Cho:Cr ratios in the DS and control neonates varied based on the TE of the acquisition. The Cho:Cr (TE 55 ms) ratio, was not to differ between the two groups (p =0.12); however, Cho:Cr (TE 144 ms) ratio was lowered in DS neonates (p < 0.05).

(A) Myo-inositol:Choline (TE 55ms), (B) Myo-inositol:Creatine (TE 55ms), (C) N-acetylaspartate:Choline (TE 144ms), (D) N-acetylaspartate:Creatine (TE 144ms), (E) Choline:Creatine (TE 55ms), and (F) Choline:Creatine (TE 144ms). Post-menstrual age (PMA).
No significant differences were found in metabolite ratios between male and female neonates in either the control or DS groups. We could not analyse the differences in metabolite ratios in DS participants with (n=14) and without (n=4) a congenital heart defect (CHD), due to insufficient numbers for comparison. Neither, DS neonates without a cardiac defect (n=4), nor those born prematurely (prior to 37 weeks, n=6) were outliers.
Fetal Mass spectrometry study
We measured the total amount of the metabolites Creatine, Choline, Myo-inositol and NAA (Figure 4; Table 6) in cortical tissue from fetal brain. 14 fetuses with DS (median (range) - 13.50 (10-19) PCW and 30 control fetuses (15.00 PCW, (10-20) PCW) were used in this study. We found a statistically significant elevation in total Myo-inositol in DS fetal brains compared to controls (p < 0.05, d = -0.7), when controlled for age (Figure 4C). This difference appears to be greater in the 10-12 pcw samples analysed. No group differences were observed in Creatine, Choline and NAA. However, the trajectories over gestation appear to be lower in DS as compared to controls. Specifically, Myo-inositol and Creatine appears to be on downward trajectories in DS, as compared to the control population. For comparison with the MRS study, ratios were calculated for the mass spectrometry cohort, from the absolute values (Supp. table 1). The ratios for Myo:Cho and Myo:Cr were found to be significantly different between the two groups (p<0.05), supporting our MRS findings. However, a statistically significant difference was not detected for the other ratios.
LME results, Cohen’s d values and effect sizes for the DS vs control cohort – Mass spec data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Lines indicate mean.
Discussion
To the best of our knowledge, this is the first study to report elevations in both, absolute total myo-inositol values ex vivo in the human fetal brain with DS, and in vivo MRS brain data in neonates with DS. In this study, we investigated metabolic profiles in the developing brain with DS, with focus on Myo-ins. We found significant alterations in the metabolite ratios of Myo-ins and NAA in the brain of neonates with DS compared to typically developing neonates. We also found that absolute Myo-ins was elevated in fetal brain tissue in DS, prior to 20 weeks when compared to controls, markedly so earlier in gestation at 10 weeks.
Our study found Myo-ins ratios were significantly higher in the DS neonates, compared to the control population in the deep gray matter. In support of an elevation of Myo-ins in DS, our second study assessing fetal brain tissue with mass spectrometry also demonstrated elevated Myo-ins levels in the cortex, when compared to controls prior to 20 weeks gestation, particularly in the earliest ages examined, 10-12 pcw. Thus far, increased absolute Myo-ins levels have not been reported in the human developing fetal DS brain using mass spectrometry. As the early imbalances in Myo-inositol levels, from 10-12 pcw, may account for the overall observed increase in DS cases, a larger sample size would be needed to determine if there is a consistent, comparative elevation in absolute myo-ins concentration in DS fetuses.
Myo-ins is specifically relevant in the pathology of DS, as the NA+/Myo-inositol co-transporter (SLC5A3) is located on HSA21 (Berry, Wang and Dreha, 1999). Inositol transporters such as sodium-Myoinositol cotransporter-1 (SMIT1), which are encoded by the SLC5A3 gene, are responsible for regulating brain Myo-ins levels, by co-transporting two sodium ions along the concentration gradient to aid active transport of Myo-ins across cell membranes (Fenili et al., 2011; Dai et al., 2016). Myo-ins has been shown to be increased in trisomic cortical neurons (40% compared to diploid), generated from a mouse model trisomic for mouse chromosome 16 (which shows large synteny with HSA21) (Acevedo et al., 1997). Elevated levels of Myo-ins have also been found in amniotic fluid of human DS pregnancies (15-18 weeks GA) (Santamaria et al., 2014). SMIT1 is overexpressed in human DS and associated with increased intracellular levels of Myo-ins in adults (Shonk and Ross, 1995; Cárdenas et al., 2017). Given the association of Myo-ins transporters and HSA21, our results, likely reflect an elevation in incorporated intracellular Myo-ins concentration.
Myo-ins is a glial cell marker and increased levels have been associated with neuroinflammation (Chang et al., 2013). It is speculated that abnormal Myo-ins metabolism may be associated with the initial cognitive impairment in DS as well as being a predisposing factor to AD. Increased concentration of Myo-ins in AD and mild cognitive impairment (without AD) have been supported by histopathologic findings of demyelination and increased gliosis, which is thought to be related to a combination of disintegration of white matter fibres, microglial activation or astrogliosis (Siger et al., 2009). There is increasing evidence from histological studies from mouse models of AD, supporting a link between elevated concentrations of Myo-ins and astrocytic activation (evidenced by an increase in glial fibrillary acidic protein (GFAP) staining), in response to inflammation (Harris et al., 2015).
Increased Myo-ins:Cr have been identified using MRS, in both grey and white matter, in DS patients (Shonk and Ross, 1995) and non-DS patients with AD and dementia, and found to precede associated decreases in NAA, atrophy, neuronal loss and cognitive impairment (Öz et al., 2014). Additionally, higher hippocampal concentrations of Myo-ins have been reported in DS adults without dementia, compared to healthy controls (Beacher et al., 2005). Dietary intake has been shown to directly influence plasma levels of Myo-ins, but not those in the CSF, which is thought to be a consequence of global dysregulation in metabolism or of increased dosage of a single gene on HSA21 (Holub, 1986; Shetty et al., 1995). Shetty, et al., (1995) placed DS (age: 22-63 years) and control (age: 23-69 years) participants on a low monoaminergic diet for 72 hours, prior to collecting samples from a lumbar puncture. They reported a 30-50% increase in Myo-ins concentrations in the CSF of DS subjects compared to controls (Shetty et al., 1995; Fisher, Novak and Agranoff, 2002). As highlighted, the majority of Myo-ins studies have investigated levels in the adult brain, as such, our findings of elevated concentrations in the fetal brain are particularly interesting for further research and as a target for potential interventions.
The association of elevated Myo-ins levels with cognitive impairment in the adult brain, led to a recent clinical trial which aimed to reduce elevated brain Myo-ins levels in adults with DS with scyllo-inositol but found no apparent behavioural or cognitive improvements (Rafii et al., 2018). The authors attribute the result to small participant numbers, short trial duration and the potential practice effects on exploratory cognitive testing. Mouse studies of DS using the Ts65Dn model, have shown promising results in successfully reducing levels of Myo-ins with lithium treatment in the adult mouse, rescuing both synaptic plasticity and memory deficits (Huang et al., 1999; Contestabile et al., 2013). Lithium has been presented as potential treatment option in children with mood disorders, as well as those with intellectual disorders (Yuan et al., 2018). Clinical trials of lithium in children with intellectual disabilities reported positive effects on cognition with only mild, reversible side effects (Yuan et al., 2018), highlighting a potential avenue of research and prospective treatment for DS children with elevated Myo-ins.
In this MRS study we also observed elevated NAA:Cho and NAA:Cr ratios in DS compared to controls. However, this was not apparent prior to 20 weeks gestation, where we saw no difference in absolute NAA values in the fetal brain. Our MRS study also found increasing NAA:Cho and NAA:Cr ratios, with advancing PMA in both our DS and control cohorts. Previous studies in a typically developing fetal cohort (n=35; 30-41 weeks GA) have shown a similar increase in NAA:Cho and NAA:Cr ratios, increasing GA, coupled with a decrease in Cho:Cr (Kok et al., 2002). NAA is one of the most concentrated molecules in the brain. Increasing levels of NAA are reflective of brain maturation and attributable to dendritic and synaptic development and oligodendroglial proliferation and differentiation (Kato et al., 1997; Battin and Rutherford, 2002). Previous studies in adults with DS have reported lower NAA ratios (Śmigielska-Kuzia et al., 2010; Lin et al., 2016). This is thought to be an early marker of neurodegenerative changes, preceding structural changes as visualised by MRI. This is the first evidence of elevated absolute NAA in the early developing brain in DS. The significance of this result is not yet clear and may reflect abnormal acceleration of development and premature exhaustion as a consequence of potential mitochondrial dysfunction (Coskun and Busciglio, 2012)
In this study Cho:Cr ratios were found to decrease with increasing PMA at scan in both the control and DS cohorts. Total Cho and Cr in the fetal brain was not found to differ prior to 20 weeks in the DS and control tissue. Cho plays a role in lipid membrane synthesis and degradation, and in myelination, and is found in both glia and neurons (Kok et al., 2001; Girard et al., 2006). Adults MRS studies in a healthy ageing population found that Cr levels elevated over time, were thought to be a marker of decreased brain energy metabolism and associated with ageing-related mild cognitive impairment, or dementia in more extreme cases (Ferguson et al., 2002). We observed a significant decrease in Cho:Cr in our DS cohort compared to controls, at TE 144ms, but not at TE 55ms. A potential reason for this may be the limited number of cases at TE 55ms between 40-44 weeks PMA. This finding is unlikely to be associated with advanced myelination and maturation of the DS brain, as previous studies have observed delayed myelination in post-mortem fetuses and neonates (Wisniewski and Schmidt-Sidor, 1989). Given that the results from this study show a decrease in the Cho:Cr ratio, it is reasonable to propose that a decrease in choline may be responsible, but perhaps less likely, for the unexpected increase in NAA:Cho ratio.
To assess whether alterations in metabolite values exist and could be measured in utero, we performed a preliminary fetal MRS study on a small cohort of fetal DS cases (n=11) (Patkee et al., 2020), compared with age-matched controls (n=36). Fetal DS data was acquired at a combination of 1.5T and 3T field strengths, whilst the control data was exclusively acquired on a 1.5T scanner. Data was analysed using the same methodology as the neonatal cohort. Unfortunately, much of the data was found to be heavily motion artefacted and corrupted, with only a few cases which had interpretable spectra, therefore the results were deemed to be inconclusive. This is a limitation of the wider study, as we are only able to comment on fresh-frozen fetal tissue, as in vivo fetal MRS is thus far unreliable. Future work will aim to optimise MRS sequences in the fetus to account for motion and to acquire a greater volume of data, to account for the corrupted spectral acquisitions.
Limitations
Ideally, we would aim to acquire MR spectra at both echo times and from more than one region of interest. However, due to technical challenges, such as excessive motion or a neonate becoming unsettled during a scan, it was not always possible to acquire adequate quality MRS at both echo times in all cases. Shorter TEs yield a much better SNR for the detection of metabolites, however at these TEs lipid signal contamination can be found. Longer TEs produce spectra with a much clearer baseline and a reduction in lipid signals which allows for easier metabolite quantification. Due to the intrinsic variation in T2 relaxation times across the metabolites between short and long TEs, normal spectra heights may/will vary when measured at different TEs. It is evident that both cohorts have small sample sizes for spectral analysis. However, we detected significant differences in the Myo:Cho, Myo:Cr, NAA:Cho, NAA:Cr and Cho:Cr (TE136 ms) ratios between DS and control neonates. These novel changes have not been previously reported in the developing brain this early in development. There are several limitations linked to the nature of working with human fetal tissue samples, that we have taken steps to address in our experimental design. Firstly, whilst a wide range of gestational ages were sampled, wherever possible three individual samples were used per age for the controls. Secondly, in both DS and control the cortical tissue used for the mass spectrometry analysis was selected randomly, to avoid bias in the cortical areas analysed. Thirdly, due to the ages of the samples collected, the majority were collected from surgical, not medical, termination procedures, reducing the effect on tissue metabolites. Finally, once analysed by mass spectrometry, all readings were corrected for cortical tissue weight and DS samples were compared to the age-matched controls.
Conclusions
In conclusion, we found significantly elevated Myo-Ins in the DS fetal brain, and elevated ratios of Myo:Cho, Myo:Cr, NAA:Cho and NAA:Cr in the developing neonatal brain with DS, compared to a typically developing population. DS is a multifactorial disorder with a wide spectrum of phenotypes attributable to a significant degree of individual variability in genotypes (Karmiloff-Smith et al., 2016; Baburamani et al., 2019). The differences in metabolite ratios may reflect ongoing metabolic deviations that may underlie some of the altered brain volumes previously reported (Patkee et al., 2020) in DS and may be correlated with subsequent cognitive delay and potentially with the development of dementia in later life. Future studies will explore possible correlations with childhood neurodevelopmental assessments in this cohort to ascertain whether metabolite levels predict the degree of early neurocognitive impairment in DS.
Funding
This work was supported by the Medical Research Council [MR/K006355/1 and MR/LO11530/1]; Rosetrees Trust [A1563], Sparks and Great Ormond Street Hospital Children’s Charity [V5318]. We also gratefully acknowledge financial support from the Wellcome/EPSRC Centre for Medical Engineering [WT 203148/Z/16/Z], the Medical Research Council [MR/S025065/1] the National Institute for Health Research (NIHR) Biomedical Research Centre (BRC) based at Guy’s and St Thomas’ NHS Foundation Trust and King’s College London and supported by the NIHR Clinical Research Facility (CRF) at Guy’s and St Thomas’. The Brain Imaging in Babies (BIBS) team additionally acknowledge support from EU-AIMS – a European Innovative Medicines Initiative; and infrastructure support from the National Institute for Health Research (NIHR) Mental Health Biomedical Research Centre (BRC) at South London and Maudsley NHS Foundation Trust and King’s College London.
The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR or the Department of Health.
Competing Interests
The authors declare that they have no competing interests.
Supplementary materials
LME results, Cohen’s d values and effect sizes for the DS vs control cohort mass spectrometry ratios, as calculated from the absolute values
Acknowledgements
We thank the parents and children who participated in this study. The authors gratefully acknowledge staff from the Centre for the Developing Brain at King’s College London and the Neonatal Intensive Care Unit at St. Thomas’ Hospital; particularly the research radiologists, radiographers, clinicians, neonatal nurses, midwives and the administrative teams. In addition, we wish to thank all our obstetric and fetal medicine colleagues from our patient identification sites who have referred participants to us. We would also like to thank Dianne Gerrelli, Steve Lisgo, Berta Crespo and their teams at the HDBR for their invaluable support, and Anna Caldwell, John Halket and their team at the Mass Spectrometry facility at King’s College London for their expertise and support.
References




