

Abstract
Context The trend to delay pregnancy in the United States has resulted in the number of advanced maternal age (AMA) pregnancies to also increase. In humans, AMA is associated with a variety of pregnancy-related pathologies such as preeclampsia (PE). While AMA is known to be a factor which contributes to the development of pregnancy-induced diseases, the molecular and cellular mechanisms giving rise to this phenomenon are still very limited. This is due in part to lack of a pre-clinical model which has physiologic relevance to human pregnancy while also allowing control of environmental and genetic variability inherent in human studies.
Objective To determine potential physiologic relevance of the vervet/African green monkey (Chlorocebus aethiops sabaeus) as a pre-clinical model to study the effects of AMA on adaptations to pregnancy.
Design Thirteen age-diverse pregnant vervet monkeys (3-16 y.o.) were utilized to measure third trimester blood pressure (BP), complete blood count, iron measurements and hormone levels.
Results Significant associations were observed between third trimester diastolic BP and maternal age. Furthermore, the presence of leukocytosis with enhanced circulating neutrophils was observed in AMA mothers compared to younger mothers. Moreover, we observed a negative relationship between maternal age and estradiol, progesterone and cortisol levels. Finally, offspring born to AMA mothers displayed a postnatal growth retardation phenotype.
Conclusions These studies demonstrate physiologic impairment in the adaptation to pregnancy in AMA vervet/African green monkeys. Our data indicate the vervet/African green monkey may serve as a useful pre-clinical model and tool for deciphering pathological mediators of maternal disease in AMA pregnancy.
Introduction
Health quality and outcomes for pregnant mothers in the United States are not improving, even with the advancement of modern medicine. In fact, US pregnancy-related maternal mortalities rose 26.6% between 2000 and 20141. Moreover, while the US infant mortality rate is not increasing, it is significantly higher than that of other developed countries2. This data highlights a pressing need to understand maternal adaptations to pregnancy in an effort to improve health outcomes for both the mother and child.
Over the last several decades, women and their partners more frequently choose to delay childbirth. The reasons for this change are multi-factorial, but include educational pursuit, access to reliable contraception, and economic uncertainty3. While the overall national fertility rate has steadily declined to the lowest numbers recorded in 32 years, the rate of advanced maternal age (AMA) pregnancies, defined as 35 years and older, has risen dramatically4. From 2000 to 2014, birth rates for women under 20 declined 42% while the number of women having their first child at age 35 or older rose 23%5. The emerging trend of AMA pregnancies is paramount to understand as AMA has been associated with increased risk of several adverse maternal and fetal outcomes6–9. For example, AMA is associated with increased risk of gestational diabetes mellitus, placenta previa, and postpartum hemorrhage7. In addition, several adverse cardiovascular phenomena have been associated with AMA, including higher risk of developing hypertension and arrhythmias during pregnancy10. These conditions are clinically significant considering that 26% of pregnancy-related deaths between 2006 and 2013 had cardiovascular etiologies10, 11. Hypertension during pregnancy can also be used to predict future changes for both mother and fetus; women diagnosed with pregnancy-related hypertension experience a 2-8 fold increase in risk for future hypertension,12–17while babies born to hypertensive mothers are more likely to develop cardiovascular disease themselves18–21. These human data reinforce the need to understand the biological underpinnings of AMA in an effort to improve health outcomes for both mother and child.
Despite the known connection between AMA and pregnancy-related diseases, a gap in knowledge still exists in the pathogenic drivers of this phenomenon in humans. This can somewhat be explained by lack of control over environmental conditions in human studies, along with genetic heterogeneity in human populations. Furthermore, rodent models can lack physiological relevance to reproductive biology in humans. Therefore, a preclinical model with physiological relevance to human pregnancy as well as the ability to control environmental settings is needed to better define underlying mechanisms.
Previous non-human primate (NHP) models have noted similarities between humans and NHPs in hormone physiology during pregnancy and in reproductive biology, which demonstrates their potential as appropriate human pregnancy models7. To address this pre-clinical need, we posit and describe herein the use of the vervet/African green monkey (Chlorocebus aethiops sabaeus) to model the effects of AMA on maternal adaptation to pregnancy. We demonstrate this model as a pre-clinical platform to garner mechanistic insight, in a tightly controlled environmental setting, into the effects of AMA on pregnancy-induced pathologies, with strong potential for human translational relevance. Our findings demonstrate dysregulated hormonal, cardiovascular, and immunological responses to pregnancy in AMA vervets, all modeling known maladaptive responses to pregnancy in humans. Collectively, our results show that vervets are a clinically relevant model to study the effects of AMA in both maternal and fetal aspects and allow us to compensate for the shortcomings of existing human and animal studies.
Materials and Methods
Cohort Selection
A cohort of 13 vervet/African green monkeys (Chlorocebus aethiops sabaeus) was selected from the Vervet Research Colony at Wake Forest University School of Medicine. All animals were colony-born, mother-reared, of known-age and were housed in speciestypical, matrilineal social groups. Pregnancy status and estimated gestational age was determined via ultrasound as previously described22. Modal age of first birth is 4 years old in this colony. Monkeys 3-9 years old were considered optimal maternal age, while monkeys 10 and older were considered to be AMA. In addition, the cohort included primiparous (n=6) and multiparous (n=7) mothers. None of the selected animals exhibited any other comorbidities such as diabetes or heart disease. Other elimination criteria for this study included active participation in other studies. All studies were conducted under the approval of the Institutional Animal Care and Use Committee (IACUC) at Wake Forest School of Medicine.
Diet
All animals were maintained on a standard chow diet (Monkey Diet Jumbo 5037, LabDiet, St. Louis, MO). Animals were fed ad libitum except for fasting on the day of sedated procedures.
Sedation Protocol
Animals were sedated via intramuscular injections of ketamine (10mg/kg) and midazolam (0.1mg/kg). When necessary, a booster dose (50% of induction dose) was administered to maintain sedation.
Blood pressure
Systolic and diastolic blood pressure (BP) were measured via high definition oscillometry (S+B medVET, Babenhausen, Germany) as previously desribed23, 24. Three high quality measurements were recorded and then averaged to ensure accuracy.
Complete Blood Counts
Blood was collected via femoral venipuncture into EDTA vacutainers (BD Biosciences; Warwick, RI) approximately two weeks prior to parturition and again 2-5 days postpartum; 500 μL of whole blood were isolated and sent to IDEXX laboratories (Westrbrook, ME) for analysis including a complete blood count (CBC). The remaining blood was centrifuged, and the resulting plasma was collected and stored at −80°C for further analysis.
Ultrasound
Under sedation, ultrasound (Sonosite M-Turbo; Bothell, WA) was used to measure the biparietal diameter of the fetus in utero as previously described22. Three measurements were recorded to calculate an average diameter to ensure accuracy.
Iron Assays
Plasma was analyzed with the BioVision (Milpitas, CA) Total Iron-Binding Capacity (TIBC) and Serum Iron Assay Kit (Colorimetric) according to manufacturer’s instructions. Analysis determined the unbound iron, TIBC + unbound iron, free iron and free iron + transferrin bound iron. These values were used to calculate the TIBC, plasma iron and percent transferrin saturation.
Hormone Measurements
Plasma was used to determine hormone levels via commercially available enzyme-linked immunosorbent assays for estradiol using the Estradiol Parameter Assay Kit (R&D Systems; Minneapolis, MN, USA) according to manufacturer’s instructions. Progesterone was measured with the Progesterone Human ELISA kit per manufacturer’s protocol (IBL-International; Hamburg, Germany). Finally, cortisol levels were detected utilizing a commercially available kit following manufacturer’s instructions (R&D Systems).
Statistical Analysis
When comparing two groups an unpaired student’s T-test was used to determine significance. Associations were determined with linear regression analysis. Significance was determined if p<0.05.
Results
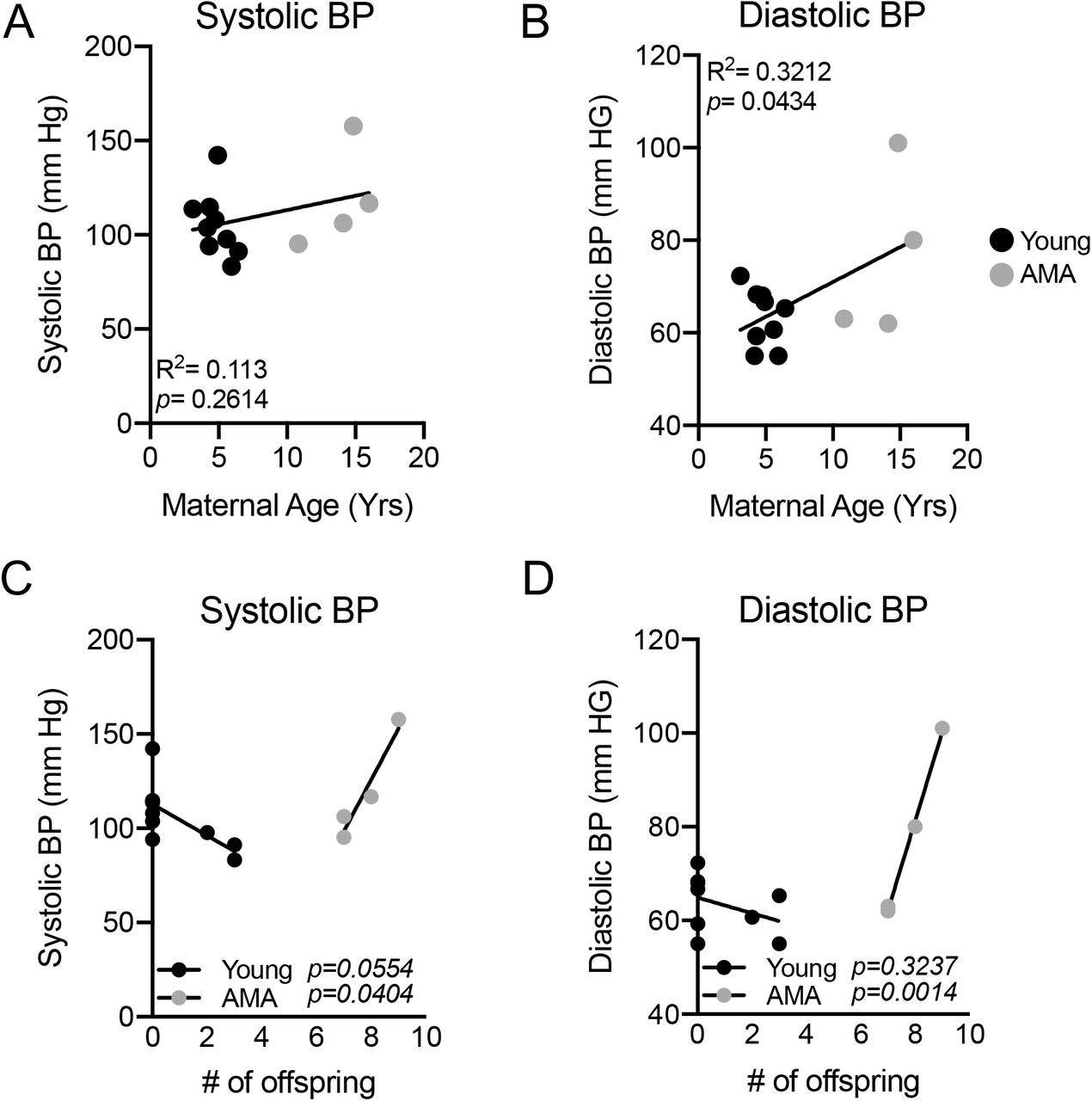
Maternal Age and Blood Pressure
Given the increased risk for the development of preeclampsia with AMA in humans25, 26, we measured BP near the end of the third trimester (approximately two weeks before parturition) in a cohort of age diverse vervets (n=13). Comparing systolic BP with maternal age revealed no significant relationship (R2=0.113; p=0.2614) (Fig. 1A). On the other hand, maternal age had a significant positive association with diastolic BP (R2=0.3212; p=0.0434) (Fig. 1B). In women, the incidence of preeclampsia decreases substantially in mothers from their first child to their second child25, 27, 28. Therefore, we wanted to determine if multiparity might mask the presence of clinical preeclampsia in our AMA cohort. There was a significant positive association between maternal age and number of off-spring (R2=0.9295; p<0.0001) (Supplemental Figure 1). Given the strong association between maternal age and number of offspring we wanted to determine if the protective effects of previous pregnancies are equivalent in young and AMA vervets. This revealed a trend for lower systolic and diastolic BP in young mothers with increasing number of pregnancies (p=0.0554 & p=0.3237 respectively) (Fig. 1C&D). Strikingly, we found in AMA a significant and strong relationship between number of offspring and both diastolic and systolic BP (p=0.0404 & p=0.0014 respectively) (Fig. 1C&D).

Linear regression between parity and maternal age. R2=0.935; p<0.0001. N=13 monkeys.

(A) Linear regression analysis between third trimester systolic BP and maternal age in vervet monkeys. R2=0.1148; p=0.2575. (B) Linear regression analysis between third trimester diastolic BP and maternal age in vervet monkeys. R2=0.3229; p=0.0428. (C) Linear regression analysis between systolic BP and # of offspring in young (black dots) versus AMA mothers (grey dots). (D) Linear regression analysis between diastolic BP and # of offspring in young (black dots) versus AMA mothers (grey dots). N=13 monkeys, 9 young mothers and 4 AMA mothers.
Leukocytosis in AMA Mothers
Activation of the maternal immune system is a well appreciated contributor to the development of preeclampsia29–31. Given the association between maternal age and increasing diastolic BP we sought to determine if maternal age altered third trimester immune cell composition. Complete blood cell counts indicated a significant positive relationship between circulating white blood cell (WBC) number and maternal age (Fig. 2A). Stratifying monkeys between young and AMA revealed significantly higher circulating WBCs in AMA mothers compared to their younger counterparts (Fig. 2B). Our initial screen to determine the cellular components contributing to leukocytosis in AMA mothers revealed no significant alterations in total circulating lymphocyte counts (R2=0.02977; p=0.5730) (Supplemental Figure 2).

Linear regression between total lymphocyte count and maternal age. R2=0.02977; p=0.5730. N=13 monkeys.

(A) Linear regression of total circulating white blood cell count and maternal age. R2=0.4085; p=0.0187. N=13 monkeys. (B) Third trimester white blood cell count in young (under 10 years of age) and advanced maternal age vervets. N=9 young mothers and 4 advanced maternal age mothers. p=0.038.
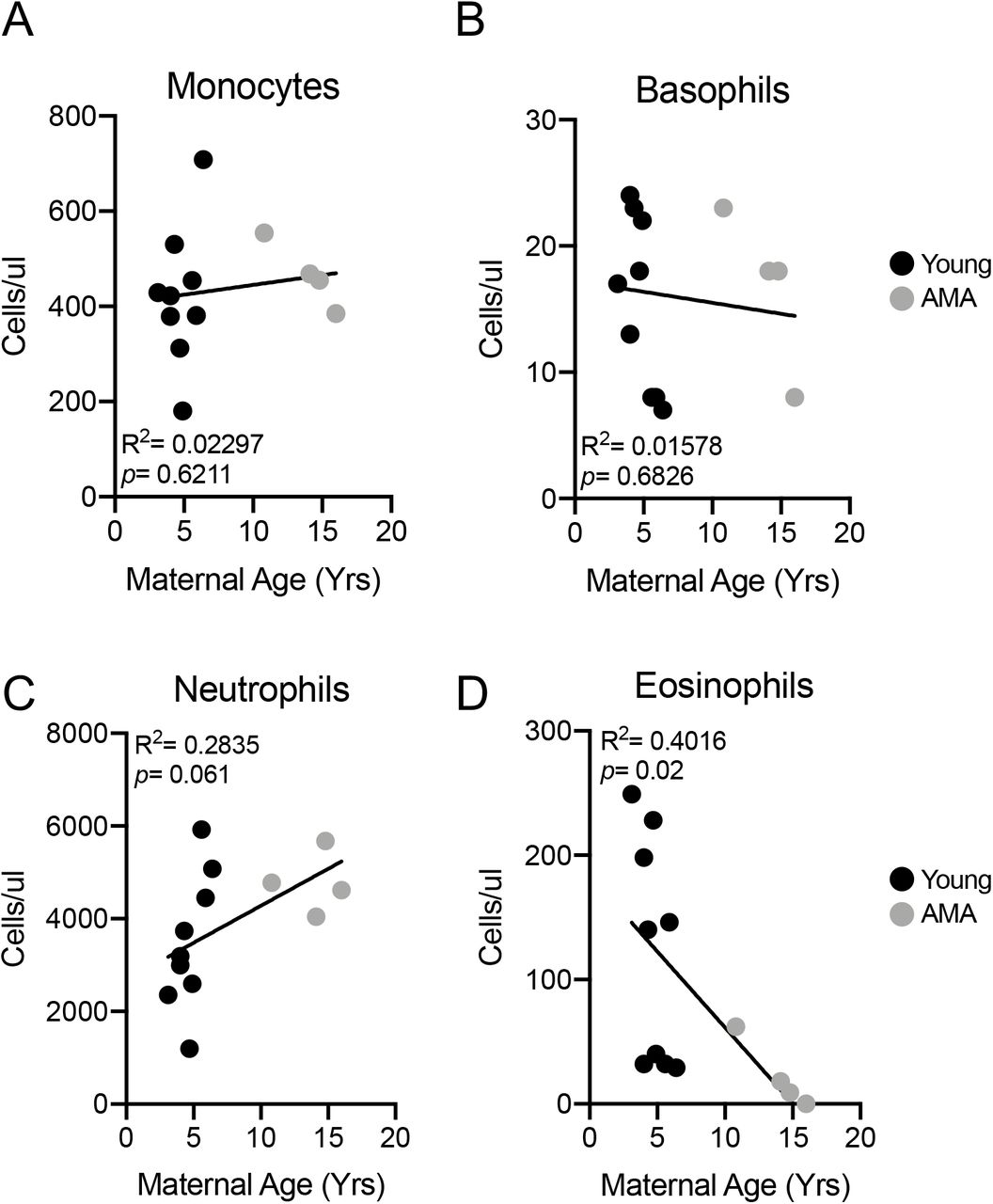
Stress Leukogram in AMA Mothers
Growing evidence indicates a role for adaptive immune cell activation in the context of preeclampsia32. We therefore assessed circulating components of the adaptive immune system including monocytes, basophils, neutrophils and eosinophils. While no alterations were observed in total monocyte (R2=0.02997; p=0.6211) and basophil numbers (R2=0.01578; p=0.6826) in the circulation related to maternal age (Fig. 3A&B), we observed trends for increased neutrophils with AMA (R2=0.2835; p=0.061) (Fig. 3C) and a significant negative association between maternal age and eosinophil numbers (R2=0.4016; p=0.02) (Fig. 3D). The presence of neutrophilia and eosinopenia is characteristic of a stress leukogram response33.
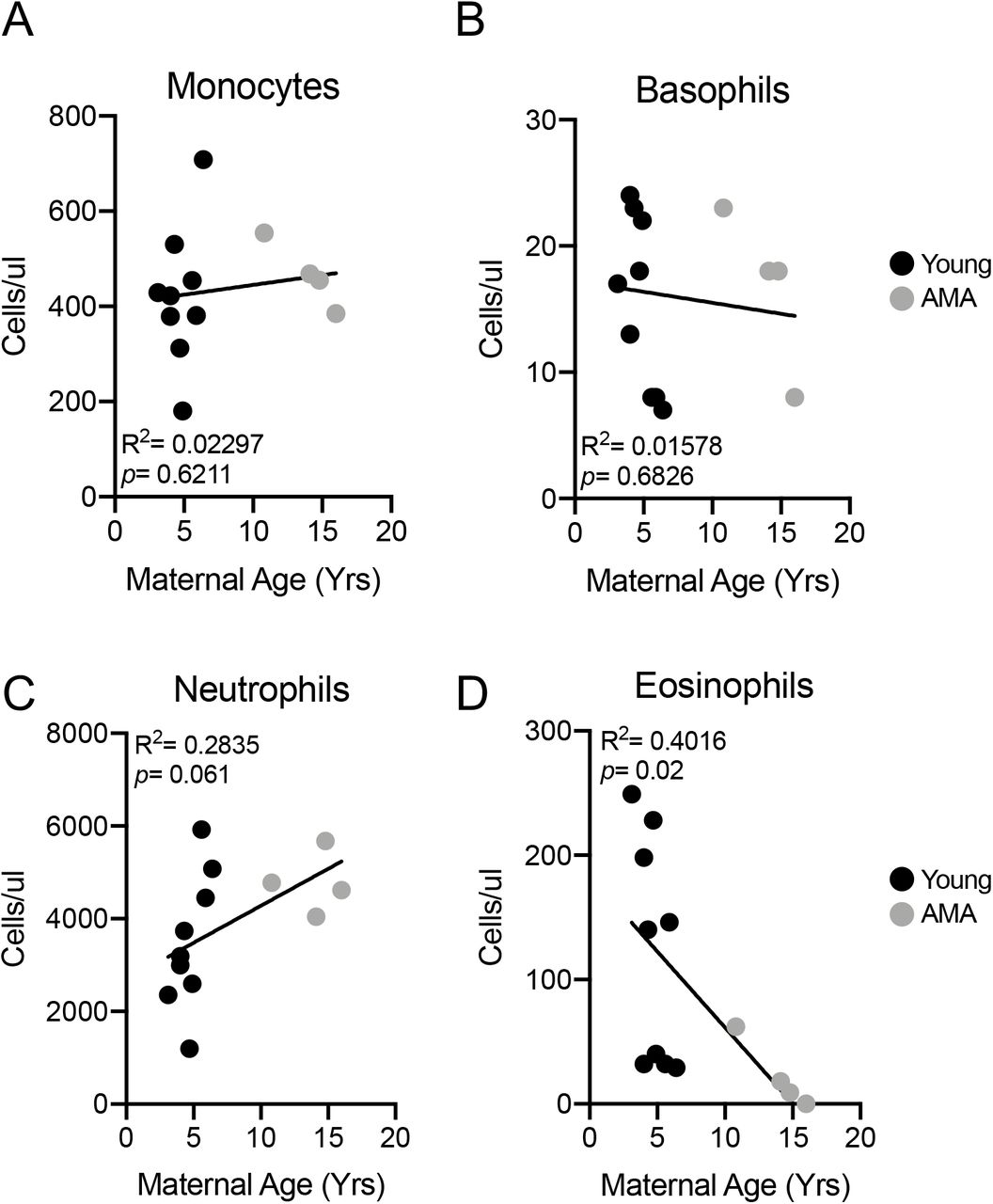
(A) Linear regression between total circulating monocyte count and maternal age. R2=0.02297; p=0.6211. (B) Linear regression between total circulating basophil count and maternal age. R2=0.01578; p=0.6826. (C) Linear regression between total circulating neutrophil count and maternal age. R2=0.2835; p=0.061. (D) Linear regression between total circulating eosinophil count and maternal age. R2=0.4016; p=0.02. N=13 monkeys.
Maternal Body Weight and AMA
To gain insight into mechanisms underlying altered immune and cardiovascular responses we assessed maternal body weight as a risk factor. We observed no significant association between maternal age and maternal pre-pregnancy body weight (Fig. 4).

(A) Linear regression between maternal body weight and maternal age. R2=0.05163; p=0.4553. N=13 monkeys.
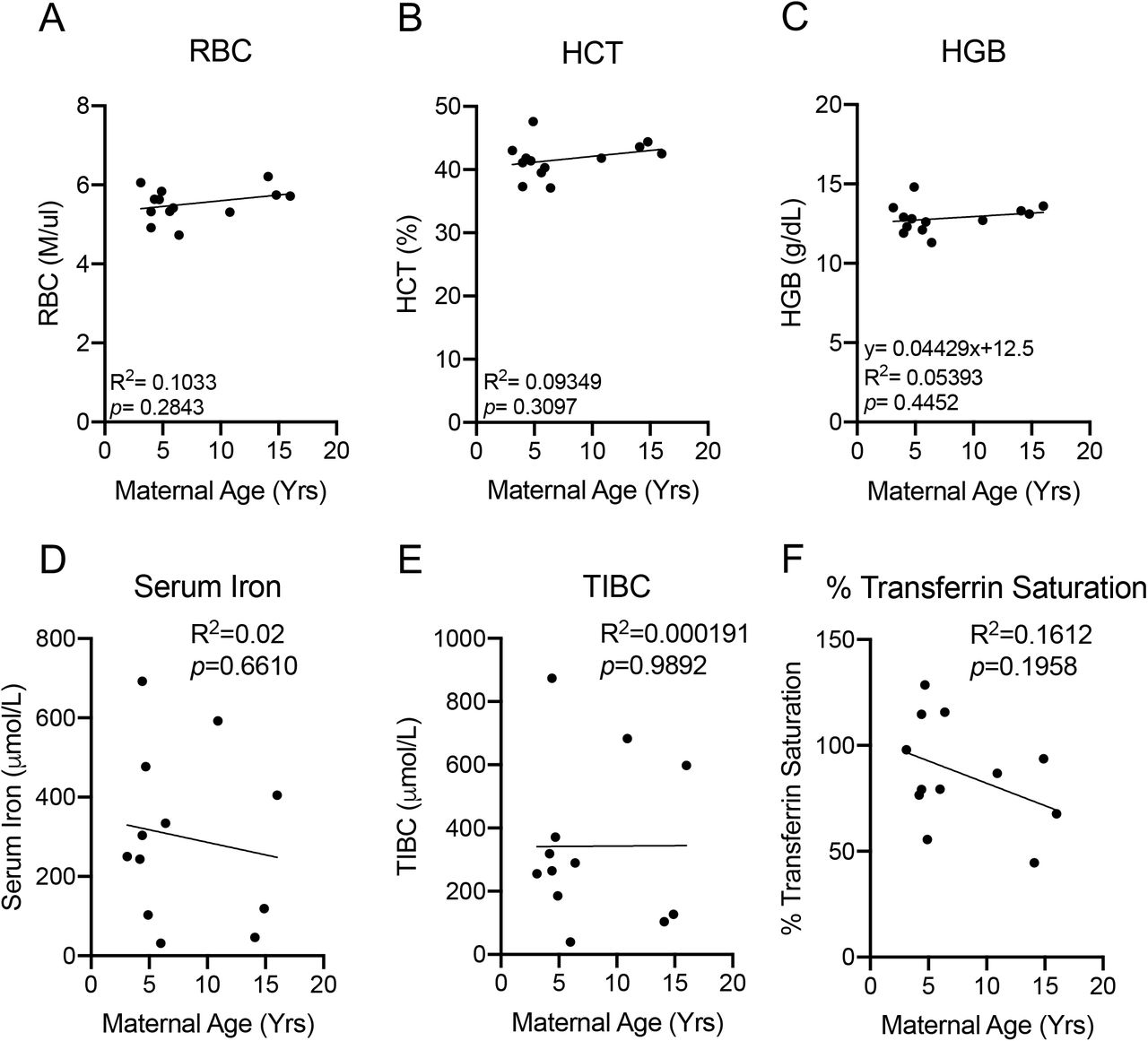
AMA does not elicit Anemia
We next determined if AMA promotes the development of gestational anemia. We evaluated several parameters associated with anemia in our cohort including red blood cell count, hematocrit and hemoglobin levels. AMA did not alter any biomarker associated with anemia (Fig. 5A-5C). Furthermore, normal serum iron levels (Fig. 5D), total iron binding capacity (Fig. 5E) and % transferrin saturation (Fig. 5F) confirmed the absence of altered iron homeostasis in older mothers.
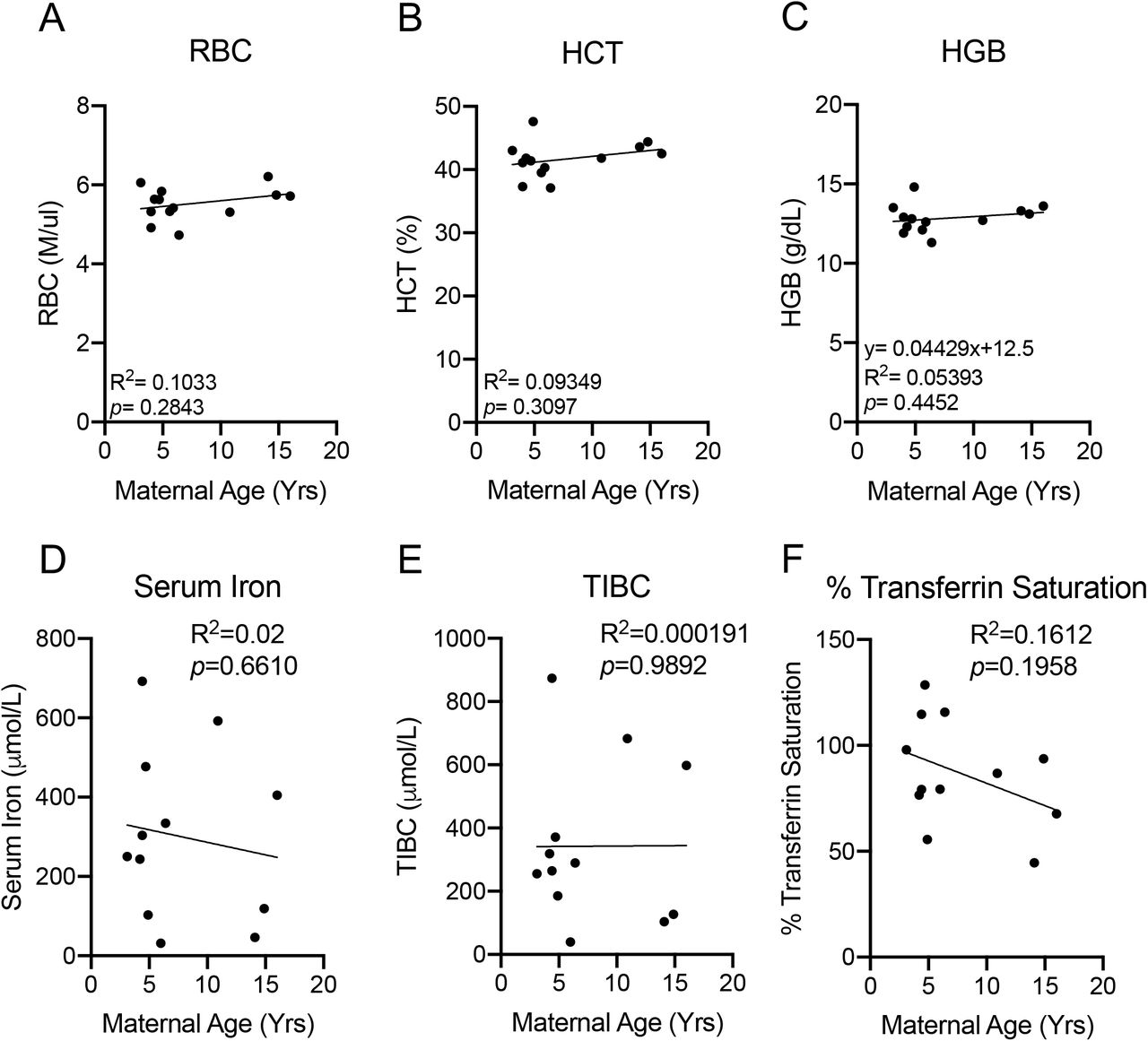
(A) Linear regression between total circulating red blood cell count and maternal age. R2=0.1033; p=0.2843. (B) Linear regression between maternal hematocrit and maternal age. R2=0.09349; p=0.3097. (C) Linear regression between maternal hemoglobin and maternal age. R2=0.05393; p=0.4452 (D) Linear regression between maternal serum iron level and maternal age. R2=0.02; p=0.6610. (E) Linear regression between total iron binding capacity and maternal age. R2=0.000191; p=0.9892. (F) Linear regression between % transferrin saturation and maternal age. R2=0.1612; p=0.1958. N=13 monkeys.
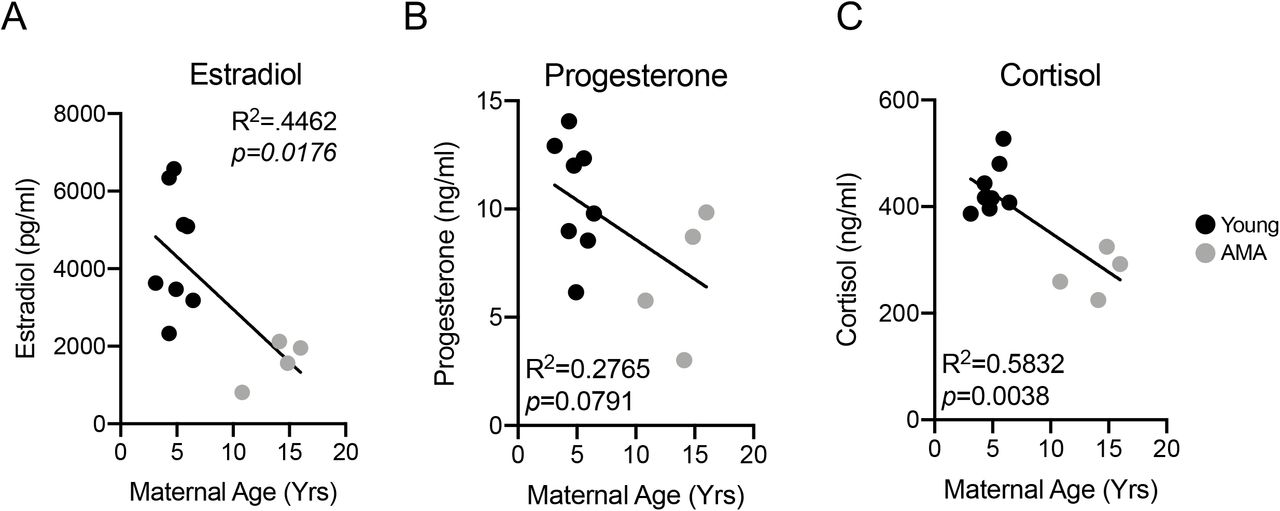
Altered Hormonal Responses in AMA Mothers
AMA is associated with low peak gestational estradiol levels34–36 and estrogen deficiency has been shown to promote diastolic dysfunction37. Therefore, we measured third trimester estradiol levels in our cohort of young and AMA vervets. Enzyme-linked immuno-sorbent assay (ELISA) revealed AMA mothers had significantly lower third trimester estradiol levels (~60% reduction; R2=0.4462; p=0.0176) (Fig. 6A). Further, we found a trend for a negative association between maternal age and circulating third trimester progesterone levels (R2=0.2765; p=0.0791) (Fig. 6B). Finally, given the presence of a stress leukogram signature in our AMA mothers, we also measured cortisol levels, revealing a significant negative relationship (R2=0.5832; p=0.0038)) between maternal age and third trimester cortisol levels (Fig. 6C).

(A) Linear regression between estradiol and maternal age. (B) Linear regression between progesterone and maternal age. (C) Linear regression between cortisol and maternal age. N=13 monkeys.
Postnatal Growth Retardation in Offspring from AMA Mothers
We measured fetal biparietal diameter approximately two weeks prior to delivery via ultrasound. No appreciable differences were observed in fetal biparietal diameter within our cohort (Fig. 7A). Accordingly, we also did not observe significant differences in infant body weights between young and AMA age mothers at birth (Fig. 7B). However, following archival growth trajectories over approximately the first year of life in a separate cohort of animals (n=28 young and n=14 aged) revealed significant growth retardation in infants born to AMA mothers (Fig. 7B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Linear regression between fetal biparietal diameter and maternal age. R2=0.03142; p=0.05624. N=13 monkeys. (B) Archival growth rates of offspring from a separate cohort of young and advanced maternal age mothers in an expanded cohort of Vervet monkeys. n=28 young and n=14 aged. * denotes p<0.05.
Discussion
AMA in humans is an established risk factor for the development of an array of pregnancy-induced pathologies6, 8, 26, 38, 39. While the relationship between maternal age and the incidence of pregnancy-related pathologies exists, pre-clinical models with similar reproductive physiology to that of humans are severely lacking. The current study clearly shows that AMA is associated with disruptions in physiological adaptations to pregnancy in vervet/African green monkeys. In particular, we found the cardiovascular system, immune system and endocrine system all display deficits in responses to pregnancy, suggesting the presence of maternal pathologies in older vervet monkeys. Additionally, first year growth trajectories were impaired in infants born to AMA mothers. These data collectively indicate the vervet monkey as a physiologically relevant pre-clinical model to study the effects of AMA on both maternal and offspring outcomes.
Human studies have revealed a selective increase in third trimester diastolic blood pressure and a decrease in systolic BP with increased maternal age40. Consistent with these findings, we observed maternal age to be significantly positively associated with diastolic BP in our vervet model. Contrary to the human studies however, we observed no relationship between age and third trimester systolic BP. These findings indicate that the vervet monkey recapitulates some, but not all aspects of altered BP regulation during pregnancy in older mothers. Gaillard et al. indicated that a woman’s maternal age per se was not consistently correlated with gestational hypertension, and that maternal body mass index might influence alterations in BP regulation during pregnancy38. In fact, maternal obesity has been shown to interact with maternal age to promote a variety of other pregnancy-induced pathologies38. We observed no association between maternal body weight and maternal age (Figure 4), which may explain differences observed between our study in vervet monkeys and human studies in the regulation of third trimester systolic BP.
Beyond elevated BP, a significant immunological component to preeclampsia exists30–32, 41–43. While leukocytosis occurs during normal pregnancy44, exaggerated leukocytosis occurs in preeclamptic patients45. Our observation in the vervet monkey that AMA mothers have significantly elevated white blood cell counts coupled to the presence of diastolic hypertension are consistent with hallmarks of human preeclampsia. Intriguingly, leukocytosis present in humans with preeclampsia is due to an increase in circulating neutrophils counts45. Similar to our other data supporting physiological relevance of vervet monkeys to humans for studying the effects of AMA, the older mothers exhibited a higher degree of neutrophilia present in their third trimester compared to young mothers, potentially exacerbating a state of mild preeclampsia.
We did observe a significant positive association in our cohort between maternal age and parity. The elevated parity in our AMA could actually be providing a protective mechanism against the development of more severe preeclampsia, as this disease is more prevalent amongst primiparous mothers25, 27, 28. We observed an uncoupling of number of previous offspring and blood pressures between young and AMA mothers. Our data suggest that previous pregnancies are associated with lowered blood pressures in younger mothers; however, in AMA mothers the number of pregnancies was positively associated with both diastolic and systolic BP. These data suggest that either AMA disrupts the protective mechanisms afforded by previous pregnancies, or, that after a certain threshold of previous pregnancies the protective mechanism of parity is lost. Parity has also been associated with immunological tolerance to certain infections during pregnancy such as malaria46–48 and multiparity has been demonstrated to confer immunotolerance in rodent models of stroke49, indicating a protective role to maternal health in multiparous mothers. While not tested in the current study, further investigation into AMA primiparous third trimester physiology is warranted to determine if multiparity is protective against the development of clinical preeclampsia.
Another known risk factor for the development of preeclampsia in humans is the presence of pregnancy-induced anemia50–52. Furthermore, maternal age and parity have been shown to be associated with the presence of anemia in humans53, 54. However, we did not observe such associations between anemia and maternal age and multiparity in our study. One explanation for the lack of association between maternal age and anemia in our study is due to diet; while maternal age is associated with the development of anemia in humans, this is largely due to insufficient iron intake during pregnancy55–57. Our vervet diet has high levels of iron (230 ppm), which could potentially compensate for AMA as a risk factor.
Estradiol is a well-known cardioprotective hormone. In the non-pregnant state, low estradiol levels, such as those observed during menopause, promote the development of cardiovascular disease58, 59. Specifically, postmenopausal women are the primary clinical population diagnosed with heart failure with preserved ejection fraction (HfpEF)60–62. The cardioprotective effects of estradiol in preventing HfpEF in estrogen deficient females has been extended to nonhuman primates such as cynomolgus macaques37. In the pregnant state, low estrogen levels have been associated with preeclampsia in humans63–67. We found AMA is associated with third trimester estrogen deficiency in vervet monkeys, consistent with human data indicating maternal age is negatively correlated with low peak estradiol levels 34–36. At the molecular level, estrogens have been shown to antagonize the effect of stress hormones68–71. We have demonstrated previously that the antagonistic nature of estrogen on stress hormones is essential for appropriate adaptations to pregnancy and proper fetal development in rodents68. Our data indicate AMA disrupts the cortisol/estradiol axis through impaired estradiol production. Furthermore, the presence of a stress leukogram in AMA vervets is suggestive of aberrant stress hormone signaling in aged pregnant vervets33.
Maternal stress in humans, like AMA, underlies long-term predisposition of off-spring to disease into adulthood. This concept is known as the developmental origin of disease72. A commonality between maternal stress and AMA is they are both risk factors for the development of intrauterine growth restriction in humans and small gestational age infants6, 8, 38, 39, 73, 74. Our ultrasound data of fetal biparietal diameter revealed no association between maternal age and head size. Furthermore, infant weight at four days post-delivery was comparable between young and AMA mothers. In humans, one driver of the small gestational phenotype is pre-term delivery75–78. This may be a possible explanation for why we did not observe low birth weights in vervets, since AMA did not elicit pre-term delivery in our cohort. Beyond low birth weights, prenatal maternal stress in humans dramatically alters postnatal growth rates of offspring. Intriguingly, the offspring growth rate phenotype is dictated by timing of maternal stress, with early stress typically leading to increased growth rates and late stress promoting decreased growth rates in offspring across 21 different mammalian species79. Our results of normal infant weight but blunted postnatal growth is suggestive that AMA in vervets corroborates human data resultant of a maternal stress response late during gestation. An additional factor within the paradigm of maternal stress is maternal investment during lactation79. We did not cross foster or perform behavioral analyses in our young and AMA vervets post-delivery, therefore we cannot determine if AMA alters maternal investment during the nursing period.
Human studies limit the ability to establish disease causality. Rodent studies on the other hand allow for experimental manipulation to test mechanisms underlying disease, but their reproductive physiology is dramatically different than that of humans. Utilizing an experimental model with direct physiological relevance would allow circumvention of these hurdles. Establishing the vervet monkey as a physiologically relevant preclinical model allows for the ability to tightly regulate environmental conditions and to collect longitudinal measurements, tissues and cells currently not feasible in human studies. This model will allow for the mechanistic dissection of how maternal age promotes pregnancy-induced pathologies with high likelihood for clinical translation and the ability to impact human health.
One primary strength of our study is the establishment of a pre-clinical model with reproductive physiologic relevance to humans for studying the effects of aging on maternal health outcomes. Furthermore, the utilization of clinically relevant assays to characterize the impact of maternal age on adaptations to pregnancy is another primary strength of our study. One weakness with our study is that we focused only on third trimester physiology. It is of the utmost importance to further delineate the effects of AMA during gestation. Moreover, our studies are observational and descriptive in nature. Future studies assessing the effects of estrogen supplementation in AMA vervets on amelioration of cardiac and immunological responses to pregnancy are much needed. Finally, the study may not be powered for certain comparisons, leading to a Type II error such as maternal body weight and anemia related factors.
Our data demonstrate that AMA in vervets summarizes several maladaptive responses observed in humans, particularly dysregulation of hormonal, cardiovascular and immunological responses to pregnancy, and establishes this model for further elucidation of the mechanisms involved in the stress responses involved in maternal adaptation to pregnancy and postnatal growth retardation in humans.
Acknowledgements
The authors would like to thank M. Christina May Long and Justin Herr for assistance with the Vervet Research Colony, and Dr. Tom Register and Ms. Maryanne Post for their technical assistance with estradiol measurements. We would also like to thank the Biomarker Analytical Core of Wake Forest University Health Sciences for their assistance with cortisol and progesterone measurements.
Footnotes
The authors report no conflict of interest.
Funding Sources: NIH P40 OD010965, NIH T35 OD010946, NIH UL1-TR001420, NIH R25 HL092618
References
- [1].↵
- [2].↵
- [3].↵
- [4].↵
- [5].↵
- [6].↵
- [7].↵
- [8].↵
- [9].↵
- [10].↵
- [11].↵
- [12].↵
- [13].
- [14].
- [15].
- [16].
- [17].↵
- [18].↵
- [19].
- [20].
- [21].↵
- [22].↵
- [23].↵
- [24].↵
- [25].↵
- [26].↵
- [27].↵
- [28].↵
- [29].↵
- [30].↵
- [31].↵
- [32].↵
- [33].↵
- [34].↵
- [35].
- [36].↵
- [37].↵
- [38].↵
- [39].↵
- [40].↵
- [41].↵
- [42].
- [43].↵
- [44].↵
- [45].↵
- [46].↵
- [47].
- [48].↵
- [49].↵
- [50].↵
- [51].
- [52].↵
- [53].↵
- [54].↵
- [55].↵
- [56].
- [57].↵
- [58].↵
- [59].↵
- [60].↵
- [61].
- [62].↵
- [63].↵
- [64].
- [65].
- [66].
- [67].↵
- [68].↵
- [69].
- [70].
- [71].↵
- [72].↵
- [73].↵
- [74].↵
- [75].↵
- [76].
- [77].
- [78].↵
- [79].↵




