

Abstract
Mediator of Cell Motility 1 (MEMO1) is an ubiquitously expressed modulator of cellular responses to growth factors including FGF23 signaling, and Memo1-deficient mice share some phenotypic traits with Fgf23- or Klotho-deficient mouse models. Here, we tested whether Memo1 gene expression is regulated by calciotropic hormones or by changing the dietary mineral load.
MLO-Y4 osteocyte-like cells were cultured and treated with 1,25(OH)2-vitamin D3. Wildtype C57BL/6N mice underwent treatments with 1,25(OH)2-vitamin D3, parathyroid hormone (PTH), 17β-estradiol or vehicle. Other cohorts of C57BL/6N mice were fed diets varying in calcium or phosphate content. Expression of Memo1 and control genes was assessed by qPCR.
1,25(OH)2-vitamin D3 caused an acute decrease in Memo1 transcript levels in vitro, but not in vivo. None of the hormones tested had an influence on Memo1 transcripts, whereas the assessed control genes reacted the expected way. Dietary interventions with calcium and phosphate did not affect Memo1 transcripts but altered the chosen control genes’ expression.
We observed that Memo1 was not regulated by calciotropic hormones or change in mineral load, suggesting major differences between the regulation and physiological roles of Klotho, Fgf23 and Memo1.
Introduction
Memo1 is an evolutionary conserved protein in all kingdoms of life that has shown intracellular expression in cytoplasma and nucleus (Schlatter et al., 2012, Haenzi et al., 2014, Moor MB, 2018). A conditional and inducible knockout mouse model with postnatal deletion of exon 2 of the Memo1 gene has resulted in a syndrome of premature aging with elevated calcemia, elevated FGF23, insulin hypersensitivity and bone disease (Haenzi et al., 2014, Moor, 2018). These traits share a partial overlap with mouse models deficient in KLOTHO (Kuro-o et al., 1997) or FGF23 (Shimada et al., 2004), two proteins which have tremendously reshaped our understanding of the regulation of calcium and phosphate metabolism by the intestine, kidney and bone (Hu et al., 2013, Moor and Bonny, 2016).
In addition, evidence from cell culture experiments investigating co-localization and phosphorylation status of adaptor proteins suggested that MEMO1 protein participates in and modulates a signaling cascade involving FGF23 and the FGFR (Haenzi et al., 2014).
Serum analyses of Klotho and Fgf23-deficient mice showed excessive 1,25(OH)2-vitamin D3 levels, a finding that has been variably found in Memo1-deficient mice depending on the genetic background (Haenzi et al., 2014, Moor, 2018). The promoters of Fgf23 and Klotho both contain vitamin D response elements (Forster et al., 2011, Orfanidou et al., 2012). FGF23 secretion is increased by parathyroid hormone (PTH) (Lavi-Moshayoff et al., 2010) and 17β-estradiol (Carrillo-Lopez et al., 2009). Regarding the regulation of Memo1, a transcriptomic analysis of rat pineal glands has detected increased Memo1 transcripts upon synthetic estrogen treatment compared to controls (Deffenbacher, 2006), highlighting a potential endocrine regulation of the transcription of the Memo1 gene. Here, we therefore tested the hypothesis that Memo1 expression can be regulated by minerals or calcitropic stimuli.
Methods
Cell culture
The mouse long bone osteocyte-Y4 cell line (MLO-Y4) was kindly provided by Lynda Bonewald (Kato et al., 1997). MLO-Y4 cells were maintained in culture in minimal essential medium alpha-modified, alpha-MEM (Gibco by Life Technologies, Carlsbad, CA, USA), containing 2.5% heat-inactivated calf serum (Sigma), 2.5% heat-inactivated fetal bovine serum (Sigma), and 1% penicillin/streptomycin (Invitrogen by Life Technologies). Serum heat inactivation was carried out in water bath at 56°C for 30min. Cells were cultured on rat-tail type I collagen (Invitrogen by Life Technologies). For vitamin D stimulation, cells were kept in serum-free medium supplemented with 10nM 1,25(OH)2-vitamin D3 (Sigma) or ethanol vehicle for 24h.
Animal experiments
C57BL/6N mice were obtained from Janvier (Le Genest-Saint-Isle, France). Mice were held in a conventional animal facility with up to 6 animals per cage, and they were fed a standard laboratory chow (Kliba Nafnag TS3242; calcium 1%, phosphorus 0.65%, magnesium 0.23%, vitamin D 1600 IU/kg, vitamin A 27000 IU/kg, vitamin E 150mg/kg, protein 18.8%, crude fat 5.6%, crude fibre 3.5%, lysine 1.1%; KLIBA Kaiseraugst, Switzerland) unless specified otherwise and were kept on 12/12 (experimental) or 14/10 (breeding) light-dark cycles. All animal experimental protocols were approved by the State Veterinary Service of the Canton de Vaud, Switzerland. For all mouse studies, sample sizes were considered based on previous results in our laboratory.
Dietary interventions
For studies of mineral metabolism, specifically designed diets and hormones were used. Calcium diet challenge experiments were carried with 5 male C57BL/6N mice per condition in their home cage, all aged 12 weeks. Mice were randomly allocated to be fed modifications of KLIBA 2222 diet containing either 0.17% (low calcium diet), 0.82% (normal calcium diet) or 1.69% (high calcium diet) (KLIBA, 2222) over 7 days. All calcium diets contained phosphorus 0.8%, vitamin A 4000 IU/kg, vitamin D 1000 IU/kg, vitamin E100 mg/kg, protein 18%, crude fat 7%, lysine 14g/kg (Kliba, 2222). For a phosphate diet challenge, 3 groups of 5 male C57BL/6N mice all aged 13-14 weeks were kept in metabolic cages and randomly allocated to be fed diets containing low (0.2%), intermediate (0.8%), or high (1.5%) phosphate content over 7 days. The phosphate diets were modifications of KLIBA 2222 diet (calcium 1.2%, vitamin A 4000 IU/kg, vitamin D 1000 IU/kg, vitamin E100 mg/kg, protein 18%, crude fat 7%, lysine 14g/kg).
Hormone injections
All hormone injections were performed in the home cage of the mice after random treatment allocation of individual ear-marked mice. For 1,25(OH)2-vitamin D3 treatment, male C57BL/6N mice aged 13-15 weeks were subcutaneously injected with 2 μg/kg body weight 1,25(OH)2-vitamin D3 (Sigma D1530) in ethanol 1% in NaCl 0.9%. The dose and application route was the same as previously used in our laboratory. Control mice were injected with 1% (v/v) ethanol in NaCl 0.9%. Mice were sacrificed 6h after injection.
For parathyroid hormone treatment, male C57BL/6N mice aged 12-13 weeks were subcutaneously injected with 80 μg/kg body weight human PTH fragment 1-34 (hPTH1-34) (Sigma P3796) in NaCl 0.9% or NaCl 0.9% alone as vehicle and sacrificed 2h after injection. The dose and application route used was determined from the literature (Kramer et al., 2010).
For estradiol treatment, male C57BL/6N mice aged 16 weeks received 1 daily subcutaneous injection of 15μg 17β-Estradiol (Sigma E8875) in ethanol 0.1% (v/v) in NaCl 0.9% for 5 consecutive days and were sacrificed 4h after the last injection. The dose per body weight and the drug application route was derived from the literature, as shown Abel et al. to induce Trpv5 expression (Van Abel et al., 2002). Control mice were subcutaneously injected with 0.1% ethanol in NaCl 0.9% for 5 days.
Mouse dissection
For euthanasia, mice were intraperitoneally injected with 0.1 mg/gBW of ketamine (Ketanarkon 100 Vet., Streuli) and 0.02 mg/gBW of xylazine (Rompun, Bayer), followed by terminal exsanguination by orbital puncture under full anesthesia and /or by cervical dislocation. Organs were collected, kidneys were cut in half, and organs were snap frozen in liquid nitrogen immediately, followed by storage at −80°C until further use.
RNA extraction
RNA was extracted using TRI reagent (Applied Biosystems by Life Technologies) according to manufacturer’s instructions. RNA pellets were dried and dissolved in RNase-free H2O. RNA concentration was measured photometrically using Nanodrop (Nanodrop 2000, Thermo Fisher Scientific, Waltham, MA, USA). RNA A260/A280 ratio was assessed, and each RNA sample was visualized on a 1% agarose gel. RNA was reverse transcribed to cDNA using the PrimeScript RT reaction kit (Takara Bio Inc, Otsu, Japan). RNA input quantities per sample were 1-2 μg for bone, 500ng for kidney or 1μg of MLO-Y4 RNA. The resulting cDNA mix was diluted 2-12x depending on tissue type.
qPCR
For quantitative gene transcript expression analysis, 2uL of cDNA was used for SYBR Green qPCR (Applied Biosystems by Life Technologies) on a 7500 Fast machine (Applied Biosystems). Samples were run in triplicates in 20uL total volume for each gene, and actin or GAPDH was used for normalization. Melting curves were obtained for every run. Program settings were: 95°C during 20 seconds, 40 cycles (95°C 3 sec, 60°C 30 sec), and for melting curve stage: 95°C 15sec, 60°C 1min, rising at 1% ramp speed to 95°C (15sec), 60°C 15sec. Data were analyzed using the delta-delta CT method. Primers were ordered from Microsynth (Switzerland), and sequences are shown in Table 1. All amplified products were visualized on agarose gels.
Primers used for qPCR.
Data analysis
Data from experiments with 2 independent groups were analyzed by t-test or Mann-Whitney U test. For comparison of 3 groups, Kruskal-Wallis test was used with Bonferroni’s Multiple Comparison post-test. All statistical analyses were conducted using GraphPad PRISM 5.03. Two-sided p-values < 0.05 were considered significant.
Results
Memo1-deficient mice resemble by some traits Klotho mutant and FGF23 KO mice (Haenzi et al., 2014), and the promoters of Klotho and FGF23 harbor regulatory sequences that can be bound by vitamin D receptors (Forster et al., 2011, Orfanidou et al., 2012). For these reasons, we determined the regulation of Memo1 gene expression by minerals and calciotropic hormones. We have previously shown that MLO-Y4 osteocyte-like cell line expressed Memo1 transcripts and protein (Moor, 2018).
Memo1 is diminished by 1,25(OH)2-vitamin D3 in vitro but not in vivo
We investigated the effects of stimulation with 10nM 1,25(OH)2-vitamin D3 on gene expression in MLO-Y4 cells (Fig 1). Known vitamin D-dependent transcripts were first assessed. Transcripts of Cyp24a1 encoding a vitamin D inactivating hydroxlyase (Fig 1a) and of osteoclast regulator Rankl (Fig 1c) were increased upon 1,25(OH)2-vitamin D3 treatment, whereas transcripts of FGF23 regulator Phex and expression of bone-derived hormone osteocalcin / bone gamma-carboxyglutamate (Bglap) were diminished (Fig 1b, 1d). Memo1 transcripts were reduced by 20% upon 1,25(OH)2-vitamin D3 treatment (Fig 1e).

1,25(OH)2-vitamin D3 treatment significantly increased Cyp24a1 transcripts (A), decreased Phex transcripts (B), increased Rankl transcripts (C), decreased Bglap (osteocalcin) transcripts (D), and decreased Memo1 (E) transcripts in MLO-Y4 cells. n=6 per condition, * p<0.05 (t-test).
Next, we intraperitoneally injected 1,25(OH)2-vitamin D3 in mice. Six hours post injection we harvested the kidney and tibia of these animals and investigated the mRNA levels of Memo1. Memo1 RNA levels remained unchanged in the kidney (Fig 2a) and tibia (Fig 2b) compared to control animals injected with vehicle only. As experimental controls, we chose Cyp24a1 and Fgf23. Renal transcripts of Cyp24a1, the gene encoding the vitamin D inactivating enzyme cytochrome P450 24a1, were upregulated by 1,25(OH)2-vitamin D3 compared to vehicle (Fig 2c). In addition, expression of Fgf23 in the tibia was increased by 1,25(OH)2-vitamin D3 (Fig 2d).

Memo1 transcripts were assessed in kidney (A) and tibia (B) and were indifferent 6h after 1,25(OH)2-vitamin D3 injection, whereas renal Cyp24a1 transcripts were over 10-fold increased in comparison to vehicle control (C). Fgf23 gene expression in tibia was increased by 1,25(OH)2-vitamin D3 compared to vehicle (D). n=5 per condition, * p<0.05 (Mann-Whitney-U test).
Memo1 is not regulated by dietary calcium
Next, we determined the effect of varying dietary calcium content for 7 days on Memo1 gene expression. RNA was obtained from a previous experiment performed in our lab (366). In the kidney (Fig 3a) and in the tibia (Fig 3b) of these mice exposed to three different calcium-containing diets (0.17%, 0.82% and 1.69%), no change in Memo1 gene expression was observed. A change in gene expression of Casr encoding the calcium-sensing receptor serves as an experimental control for the dietary intervention and was reported for the samples we used in (O’Seaghdha et al., 2013).

Memo1 transcripts in kidney (A) and tibia (B) were not significantly affected by different dietary calcium intake; n=5 per diet (Kruskal-Wallis tests).
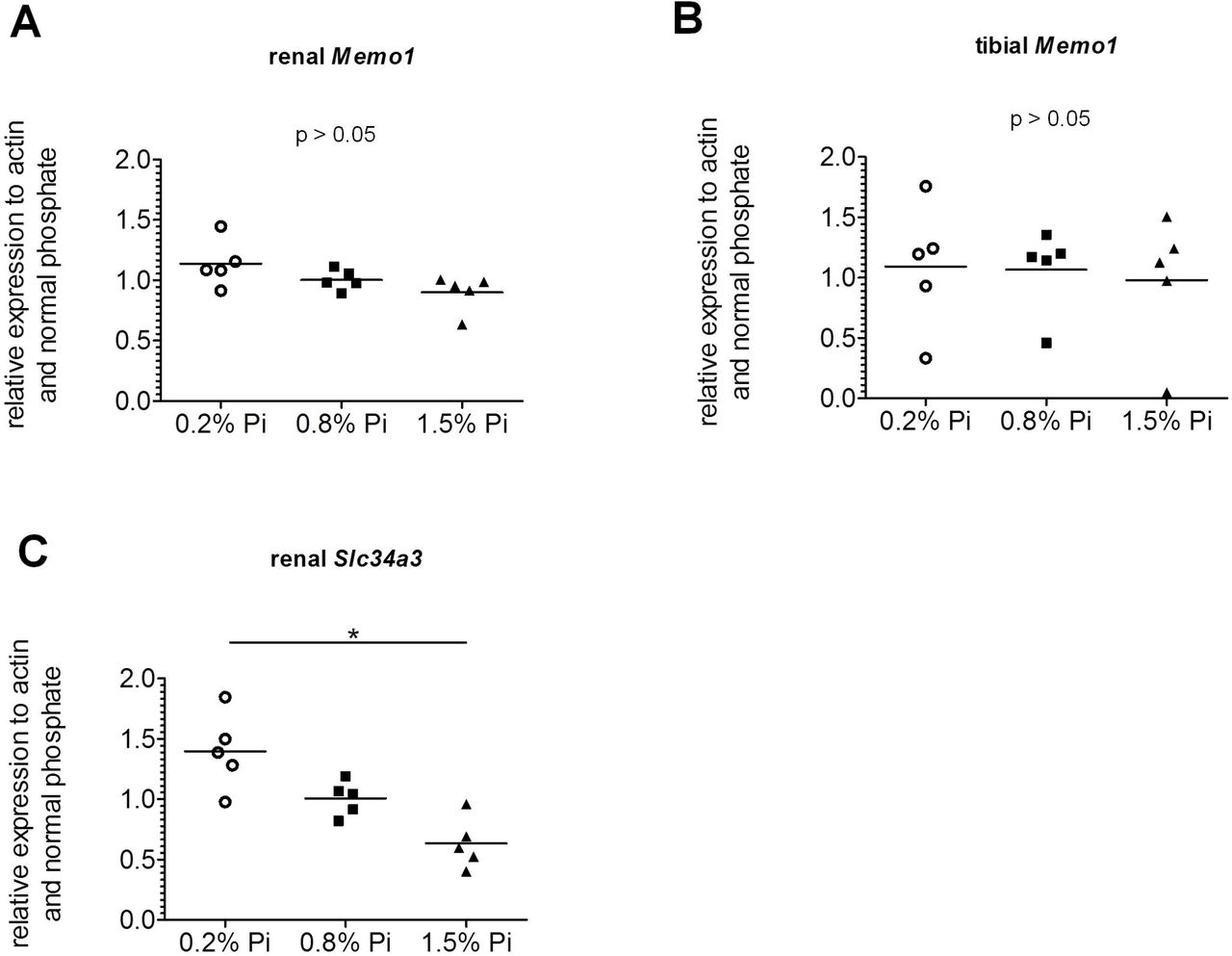
Memo1 is not regulated by dietary phosphate
We investigated the influence of different systemic phosphate loads on Memo1 expression. We showed that different dietary phosphate contents (0.2%, 0.8%, 1.5%) did not significantly affect Memo1 transcript levels in kidney (Fig 4a) or in the tibia (Fig 4b). As an experimental control gene we used renal transcripts of Slc34a3 encoding sodium-dependent phosphate transporter type 2c (NaPi2c). Renal Slc34a3 was increased, as expected, under low phosphate and decreased under high phosphate diets (Fig 4c).

Memo1 transcripts abundance in kidney (A) and in tibia (B) were not significantly changed by dietary phosphate contents. Neither bone Memo1 (B). Renal Slc34a3 transcripts were used as an experimental control gene and were affected by dietary phosphate content (C). N=5 per condition; *p<0.05 (Kruskal-Wallis test with Dunn’s multiple comparisons correction).
Memo1 unchanged by PTH
To determine the effect of PTH on Memo1, human PTH fragments 1-34 were subcutaneously injected to wildtype mice, and the animals were euthanized after 2h. Memo1 gene expression in kidney (Fig 5a) or in tibia (Fig 5b) remained unchanged upon PTH treatment. Transcripts of Cyp27b1, the gene coding for the renal vitamin D activating enzyme cytochrome P450 27b1 were increased upon PTH compared to NaCl 0.9%-treated controls (Fig 5c).
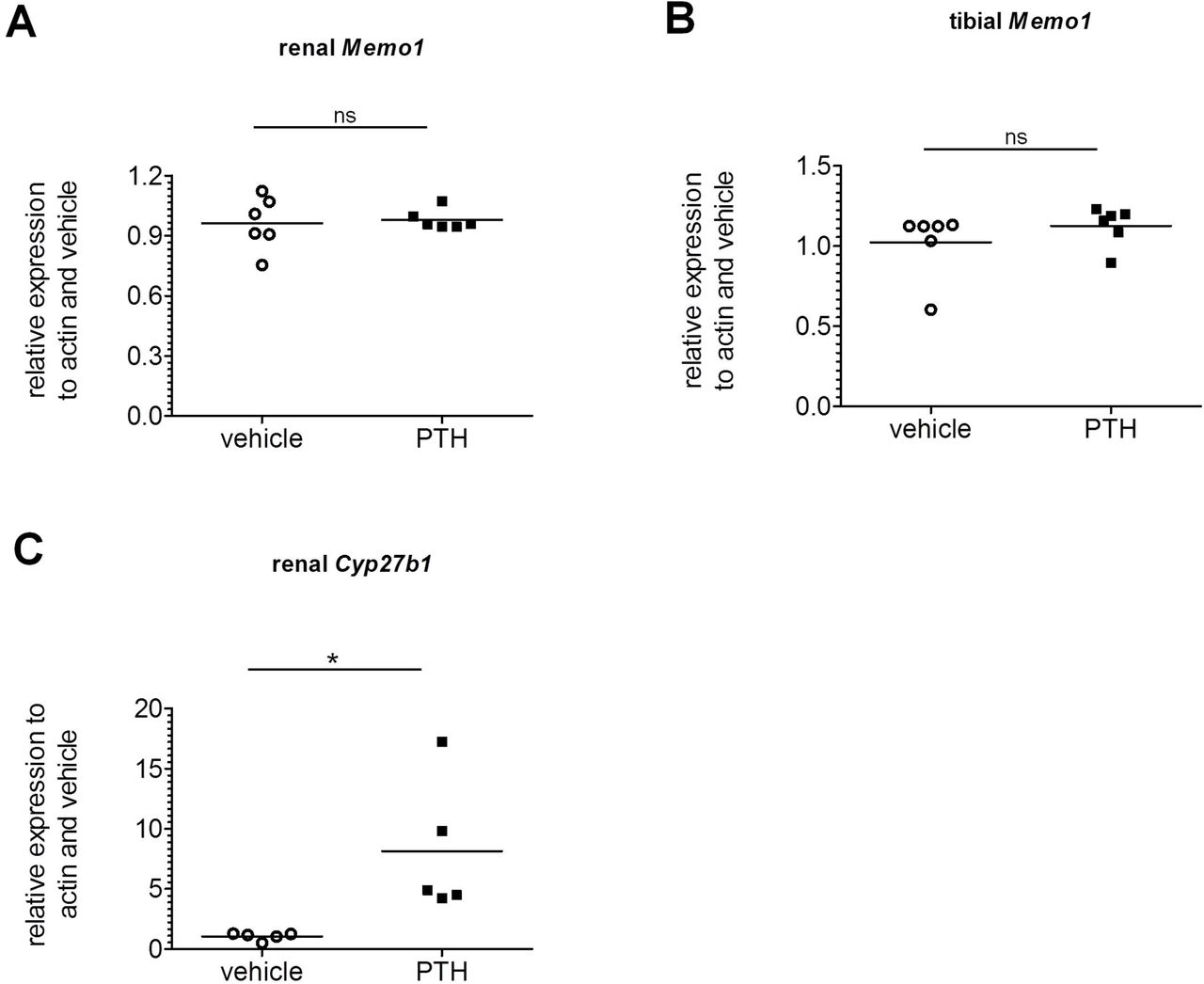
Human PTH1-34 or NaCl 0.9% vehicle was injected 2h prior to dissection, and transcripts of Memo1 in kidney (A) and tibia (B) were unchanged between experimental conditions. Transcripts of Cyp27b1 (C) were increased by PTH1-34. n= 6 per condition; ns, not significant; * p<0.05 (Mann-Whitney-U test).
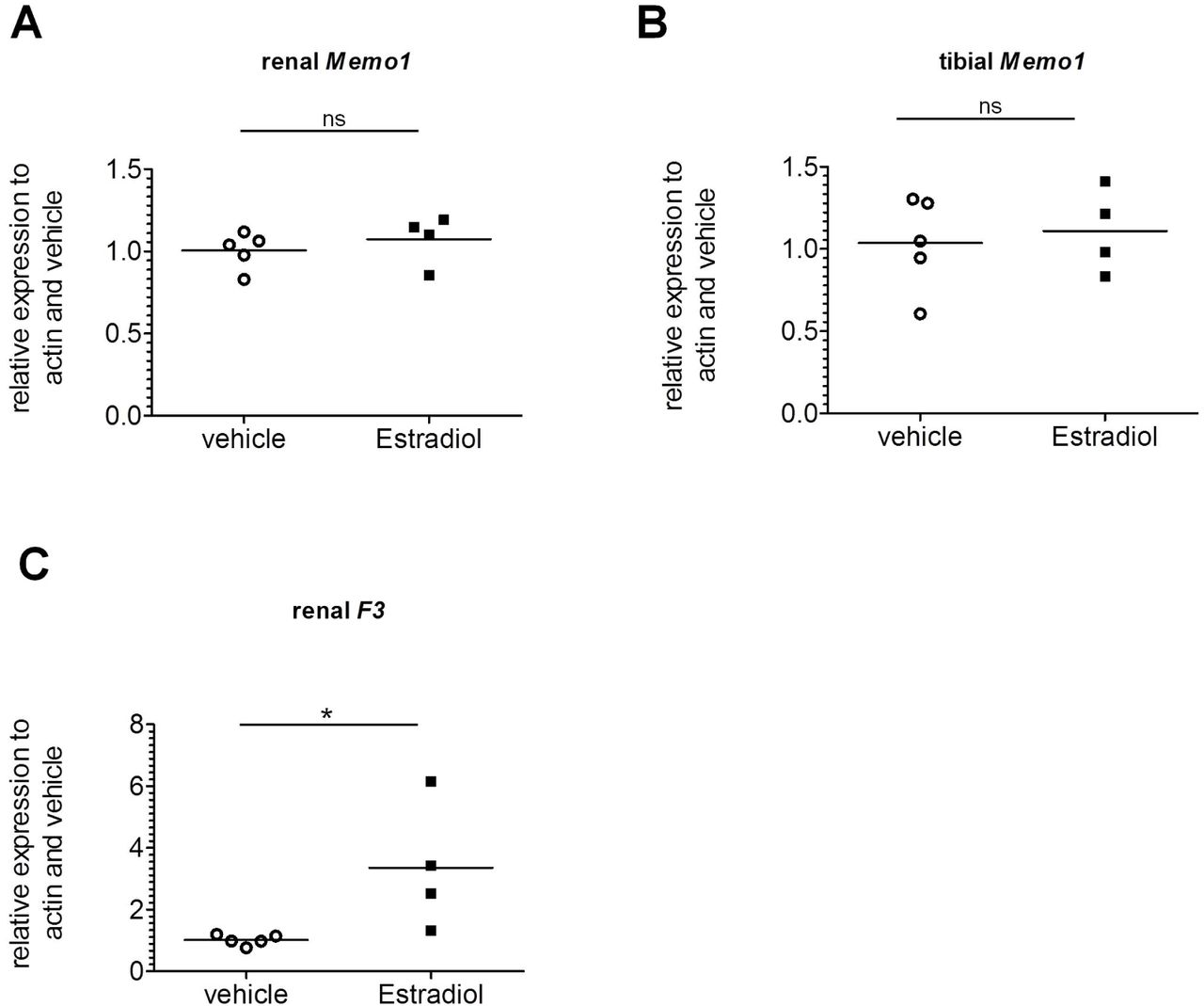
Memo1 unchanged by estradiol
As sex hormones exert effects on both renal calcium transport proteins (Van Abel et al., 2002) and FGF23 in the bone (Carrillo-Lopez et al., 2009), we tested if Memo1 is a target gene induced by estradiol. We subcutaneously injected 17β-estradiol once daily over 5 days. This induced the expression of the control gene F3 encoding coagulation factor III (Fig 6c), but gene expression of Memo1 in the kidney (Fig 6a) and in the bone (Fig 6b) both remained unchanged compared to mice injected with vehicle.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Memo1 transcripts assessed by qPCR in kidney (A) and tibia (B) were unchanged after 5 daily subcutaneous injections of 17β-estradiol compared to vehicle. Renal gene expression of tissue factor F3 was increased by 17β-estradiol (C). n=4 estradiol and n=5 control condition. ns, not significant; * p<0.05 (Mann-Whitney-U test).
To summarize, we found a small but significant decrease in Memo1 expression upon 1,25(OH)2-vitamin D3 exposure in vitro, but we failed to detect any major regulation of Memo1 transcript abundance upon mineral load or calciotropic hormone treatment in vivo.
Discussion
MEMO1 is expressed in the kidney where it plays an intrarenal role in the regulation of calcium transporters (Moor MB, 2018). In the bone, MEMO1 is expressed in all cell types (Moor, 2018), but its precise bone-specific function remains elusive. Here we tested the hypothesis whether Memo1 is regulated by key players in mineral homeostasis such as calciotropic hormones or dietary calcium or phosphate. As a readout, we chose Memo1 gene expression in an osteocyte-like cell line, and in bone and kidney tissues. For each intervention, an experimental control gene was assessed and revealed effects similar as shown before by others.
We observed that Memo1 gene expression was diminished in the osteocyte-like cells upon 1,25(OH)2-vitamin D3 treatment. However, in bone and tissues, we failed to detect any effect on Memo1 by all interventions that we studied. This shows a major difference between Memo1 and the most studied contributors to mineral homeostasis. As examples in the kidney, Type II sodium-dependent phosphate cotransporters are regulated by dietary phosphate supply (Bourgeois et al., 2013), and gene expression of Trpv5 encoding a renal calcium transport protein is controlled by 1,25(OH)2-vitamin D3 (Hoenderop et al., 2001). As examples in the bone, Fgf23 expression is stimulated by 1,25(OH)2-vitamin D3 (Liu et al., 2006) or PTH (Kawata et al., 2007), while dietary phosphate restriction or renal phosphate-wasting disorders reduce Fgf23 expression (Vervloet et al., 2011, Schlingmann et al., 2016, Ansermet et al., 2017). Even intravenous calcium loading in rats increased Fgf23 expression in bone and hormone concentrations in the serum (Shikida et al., 2018).
This study contains some limitations: The current interventions were confined to analysis of gene expression, but we did not directly assess Memo1 promoter activity using a reporter construct. Such an approach would more sensitively discriminate and would allow to identify possible response elements in the Memo1 promoter. However, we argue that a physiologically relevant regulation of Memo1 gene expression, if present, should have already been visible using the approach undertaken.
Further, we have assessed a single but physiologically reasonable time point, and only a narrow selection of tissues and cells. In addition to bone and kidney, the intestine would be another major turnover place for minerals. Memo1 expression and potential regulation in healthy intestine has not been investigated so far. In colorectal cancer cells Memo1 promoter activity is increased in response to the transcription factors Aryl hydrocarbon receptor / Aryl hydrocarbon receptor nuclear-translocator complex, indicating some intestinal disease relevance (Bogoevska et al., 2017).
Finally, as Memo is a redox enzyme with incompletely understood reaction partners and substrates (MacDonald et al., 2014), future studies assessing posttranslational regulation such as by oxidative modification of MEMO1 protein, subcellular localization, or changes in its putative enzymatic activity may be helpful to investigate a regulation of Memo1.
To conclude, besides a minor effect in bone cells stimulated with 1,25(OH)2-vitamin D3, we did not detect a major regulation of Memo1 gene expression upon minerals and calciotropic stimuli in bone and kidney, two organs relevant for mineral homeostasis. Further studies inquiring the regulation of this and similar genes may contribute to the understanding of the regulation of mineral homeostasis in health and renal and bone diseases.
Author participation
OB conceived the project. MBM and OB participated in experimental design. MBM performed experiments. MBM and OB participated in data analysis and interpretation. MBM wrote the manuscript. All authors critically read and commented on the manuscript and agreed to manuscript submission.
Acknowledgments
OB and MBM’s work was supported by the Swiss National Science Foundation through the special program NCCR Kidney.CH and by unrestricted grants from the Association pour l’Information et la Recherche sur les maladies rénales Génétiques (AIRG)-Suisse and from the Novartis Foundation. The authors are thankful to Candice Stoudmann and Finola Legrand for assisting with an experiment.
Footnotes
Disclosure: The authors declare that no conflict of interest exists.
References




