

Summary
Pharmacological interventions that target human ageing would extend individual healthspan and result in dramatic economic benefits to rapidly ageing societies worldwide. For such interventions to be contemplated they need to comprise drugs that are efficacious when given to adults and for which extensive human safety data are available. Here we show that dramatic lifespan extension can be achieved in C.elegans by targeting multiple, evolutionarily conserved ageing pathways using drugs that are already in human use. By targeting multiple synergistic ageing pathways, we are able to slow ageing rate, double lifespan and improves healthspan while minimize developmental and fitness trade-offs. Moreover, we established that there is no synergistic benefit in a daf-2 or daf-7 background, implying the involvement of the TGFβ and IGF pathways in this synergy. Employing lipidomics and transcriptomics analysis we found lipid metabolism to be affected resulting in increased monounsaturated fatty acids (MUFA) and decrease membrane peroxidation index. Our best drug combination showed a conserved lifespan extension in fruit flies. To the best of our knowledge, this is the largest lifespan effect ever reported for any adult-onset drug treatment in C. elegans. This drug-repurposing approach, using drugs already approved for humans to target multiple conserved aging pathways simultaneously, could lead to interventions that prevent age-related diseases and overall frailty in a rapidly ageing population.
Some of the most important findings in biogerontology are the evolutionarily conserved pathways that regulate lifespan1-6. In model organisms, mutations affecting these pathways can often extend lifespan by between 30% and 100%7,8. Combining different genetic mutations can result in synergistic lifespan extension9-11. By contrast, the effect of pharmacological interventions are typically much weaker, even when targeting the same pathways2,12,13, but this genetic synergy suggests a potential strategy for the design of novel pharmacological interventions by simultaneously targeting multiple evolutionarily conserved ageing pathways. To date there is little data on synergistic effects of pharmacological interventions on lifespan14. Here we report an in vivo approach to design novel multi-drug ageing interventions. We show how multi-drug interventions leverage pathway synergies to maximize effect size, while minimizing side effects and detrimental developmental tradeoffs through targeting distinct but interacting ageing pathways. Our ultimate aim was to design a purely pharmacological, adult-onset intervention with lifespan efficacy similar or better than those of canonical aging mutations did. Due to the lack of generally accepted biological markers for ageing15-17, lifespan studies are currently the only way to test the efficacy of ageing interventions18. We therefore developed our candidate drug combinations in a short-lived model organism, the nematode Caenorhabditis elegans. Using our approach, we identify two triple drug combinations that extend lifespan and healthspan to an extent greater than any previously reported pharmacological intervention in C. elegans13. Our synergistic drug combinations show effect sizes comparable to the classical ageing mutations while avoiding most of the tradeoffs associated with them19-21. We find that these interventions actually increase some markers of performance while slowing biological ageing rate. Moreover, we identified TGFβ as a key contributor and required pathway in mediating these synergistic effects. We also find that worms treated with synergistic drug combinations have higher MUFA:PUFA ratios and a decrease in membrane lipid peroxidation index. Finally, we confirm that this synergistic effect is also present in the fruit fly Drosophila melanogaster.
Results
Pathway and compound selection in C. elegans
Based on existing literature, we identified a set of well-characterized and evolutionarily conserved ageing pathways and lifespan extension mechanisms (Supplementary Table S1). We chose to target AMP activated protein kinase (AMPK), mammalian target of rapamycin (mTOR), caloric restriction (CR), C-Jun N-terminal kinases (JNK) and mitohormesis/mitochondrial metabolism as primary longevity regulatory pathways. For each pathway, we identified drugs and drug-like molecules reported to extend lifespan in at least one common model organism (nematodes, fruit flies or mice). We were interested in drugs that might eventually be tested in humans so we favored drugs with reported efficacy in mammals or that are already approved for human use. Based on these criteria, we initially selected eleven candidate drugs (Supplementary Table S1). We added allantoin to our study based on a report showing lifespan effects in C. elegans and a transcriptional analysis suggesting that its mode of action is unusually distinct from other compounds22. In order to test the lifespan effects, which can be highly sensitive to details in experimental conditions and can be variable between different laboratories19,23, we first carried out operator-blinded confirmatory lifespan studies using the previously reported optimal dose for each drug. We found that only five compounds extended lifespan reproducibly under conditions used in our laboratory (Fig. 1, Extended data Fig. 1, Supplementary Table S2). Generally, lifespan extension effects in our hands tended to be smaller compared to previous reports7,22,24-26 (Fig. 1, Extended data Fig. 1, Supplementary Table S2).

Drug treatment started at day 4 and continued until all worms died. Worms were transferred to fresh plate every day 3-4 days. (a-e) Mean and (f-j) Maximum lifespan of single drug treatments supplementary to figure 1. a and f – RAP, b and g – RIF, c and h – MET, d and i – PSORA, e and j – ALLAN (Mean ± SE). One-way ANOVA, Bonferroni multiple comparison ***P < 0.0001, **P < 0.001, *P < 0.01

Wild type N2 worms treated with different doses of a, RAP b, RIF c, MET d, PSORA and e, ALLAN have longer lifespan. f, Mean lifespan of worms treated with the optimal dose of each drug, mean ± SD. Each drug treatments resulted in a statistically significant lifespan extension at their optimal dose. **P < 0.001, ***P < 0.0001, log-rank with adjustment for multiple comparisons.
Gene expression signature of single drug treatments
To examine drug modes of action, we carried out transcriptomics analysis and determined differentially expressed genes (DEG) and pathway enrichments relative to untreated controls. Rifampicin (RIF) and rapamycin (RAP) followed by psora-4 (PSORA) caused the most extensive changes in gene expression, both at the gene and pathway level, while metformin (MET) and allantoin (ALLAN) showed comparatively smaller effects on gene expression (Fig. 2a Supplementary Table S3). While transcriptional analyses revealed significant overlap between affected genes, we also identified sets of genes that were unique to each individual compound (Fig. 2a). MET shows significant overlap with the other drugs, having the smallest unique set. Almost all genes affected by MET and PSORA were also affected by at least one other drug in the set (Fig. 2a). ALLAN causes fewer gene-expression changes overall than any of the other compounds, but 48% of the genes affected by ALLAN are unique, that is, not affected by any of the other drugs, while only 17% of the genes affected by MET are unique (Fig. 2a). Globally, effects of RIF and RAP were quite different from each other and from any of the other drugs. We confirmed this by principle component analysis (PCA) which showed that RIF, RAP and PSORA are well separated by the first three principle components. ALLAN and MET, by contrast, are more variable and very close to each other and to untreated control (Fig. 2b, Extended data Fig. 4c).





a, Circos plot illustrating gene overlap for single drugs and eat-2 mutant transcriptome. Purple lines link genes shared by multiple drugs. Blue lines link different genes which fall into the same GO term. A greater number of purple and blue links and longer dark orange arcs indicates greater overlap among the DGE and GO terms of each drug. (Minimum overlap = 3, minimum enrichment = 1.5, P value < 0.01). b, Three dimensional principle component analysis based on DGE (PCA). c, RAP+RIF and d, RIF+PSORA results in synergistic lifespan extension (P<0.0001, log-rank with adjustment for multiple comparisons). e, RAP+MET did not show further mean lifespan extension but showed further maximum lifespan extension compared to single drug treatments. f, RIF+PSORA+ALLAN and g. RAP+RIF+ALLAN showed synergistic lifespan extension (P<0.0001, log-rank with adjustment for multiple comparisons). h, Heatmap showing hierarchical clustering (distance metrics and linkage algorithms) of single drugs and synergistic combinations. n = 2000 for RNAseq and 50-100 for lifespan studies. *P<0.01, **P<0.001, ***P<0.0001
Drug synergies and lifespan extension
We next determine effect of different drug combinations on lifespan in nematodes. We began with MET and RAP due to their translational importance in humans14,27. However, RAP combined with MET at their optimal doses did not show any further benefit on lifespan (Extended data Fig. 2j,p). We carried out additional trials, exploring all possible combinations of optimal and half-optimal dose for MET and RAP (Extended data Fig. 2j-p). Combination of RAP and MET each at their half-optimal dose result in further extension in maximum lifespan (Fig. 2e, Extended data Fig. 2m,s) but did not result in further mean lifespan extension (Extended data 2p). These weak benefits of combining MET with RAP are consistent with results in mice where the existing evidence suggests that addition of MET to RAP results in some additional benefits mainly in males with only weak (if any) additional benefits in females14.

supplementary for figure 2. a-i, survival curve for all 10 possible combinations of five drugs at their respective optimal dose except RAP+MET. j-m, all possible combinations of RAP and MET at their optimal and half-optimal doses. n-p, Log-rank with proper adjustment. Mean lifespan of representative dual combinations, (Mean ± SE) q-s, maximum lifespan of representative dual combinations, (Mean ± SE). One-way ANOVA, Bonferroni multiple comparison ***P < 0.0001, **P < 0.001, *P < 0.01.
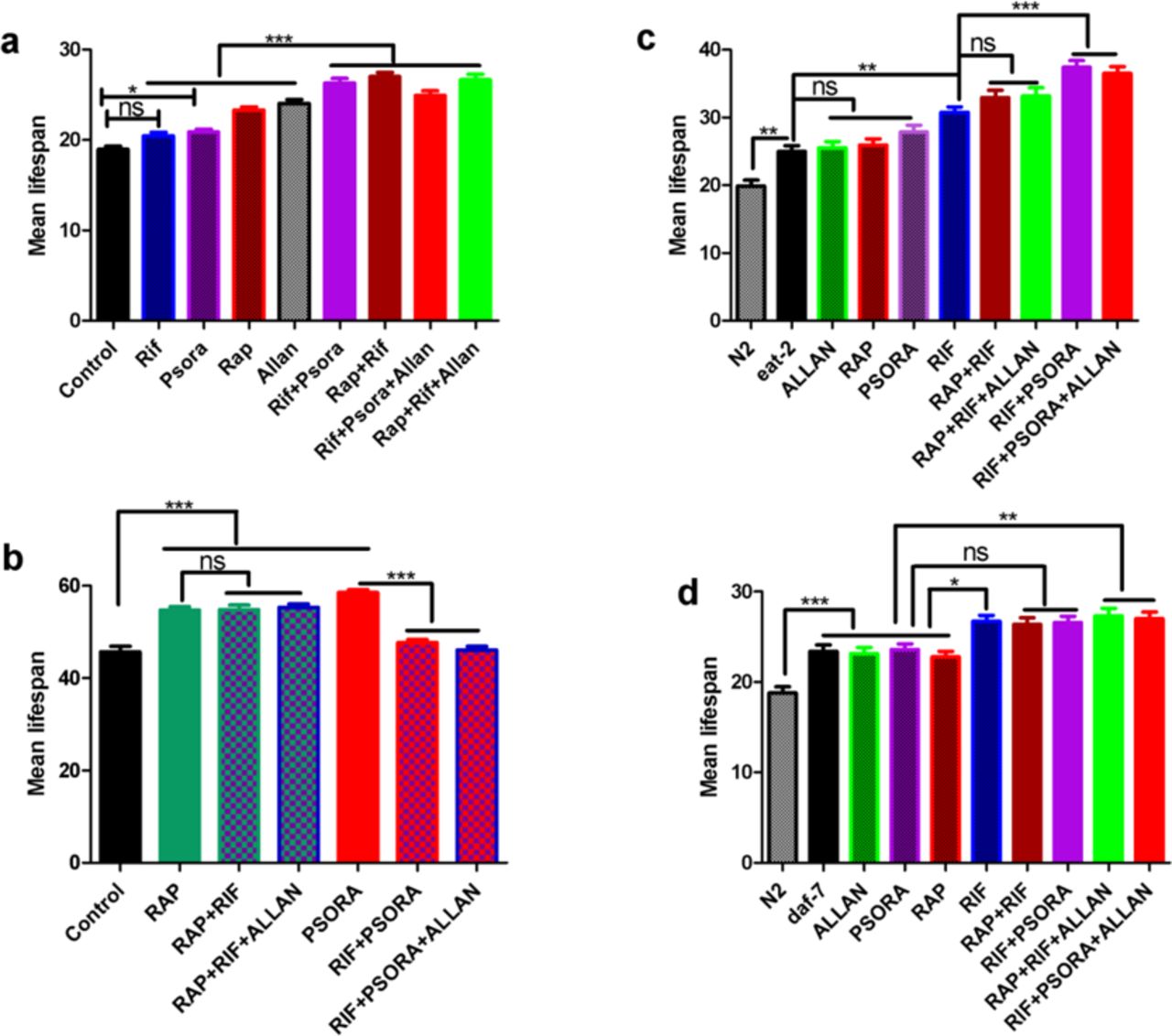
We then systematically determined the efficacy of all ten possible pairs out of the five candidate drugs (Extended data Fig. 2a-i). For these trials we define synergy based on the Higher Single Activity (HSA) model28; Drug combinations are considered synergistic if the combinatorial effect is significantly larger than any of the combination’s single drug effect at the same (ideal) concentrations as in the combination. Using this definition, we identified two synergistic combinations out of the ten pairs tested (Fig. 2c, d, Extended data Fig. 2 and Supplementary Table S2). The synergistic pairs comprise RIF with either RAP or PSORA. This means that two out of the three pairs formed by the drugs that are most well-separated in the PCA are synergistic (Fig. 2b, Extended data Fig. 4a,b). Of the remaining eight combinations, four extended lifespan as much as their best component while the other four combinations nullified each other and did not result in lifespan extension and none were toxic (Extended data Fig. 2, Supplementary Table S2).
Lifespan extension resulting from the two synergistic pairs, while larger than those of previously published drug effects, were still smaller than the benefits seen with genetic mutations9. This raises the question whether the effect size seen with the synergistic pairs represented the maximum achievable by adult-onset drug treatment or if further benefits could be gained by adding additional compounds. Since exhaustive combinatorial search of all thirty possible triple drug combinations was not feasible, we explored selected triple combinations based on the original synergistic pairs and taking into account our analysis of transcriptional changes for the single drugs. We first evaluated the combination of the three drugs best separated by PCA and involved in the two initial synergistic pairs (RAP, RIF and PSORA). However, nematodes treated with this triple combination had a shorter lifespan than seen with either of the dual combinations (Extended data Fig. 3a Supplementary Table S2). We then tested three drug combinations including ALLAN as the third component. As explained above, ALLAN has a gene expression signature that is quite distinct from the rest of our drugs. Furthermore, unlike for MET, the unique gene set of ALLAN (light orange arc in Fig. 2a) also shows no overlap at the level of GO terms (blue links) with any of the other drugs or with eat-2. This further supports the notion that ALLAN has a unique mode of action22. Therefore, we tested addition of ALLAN to the two synergistic double combinations. This addition resulted in significant additional mean and maximum lifespan extension in both cases (Figures 2f,g, Extended data Fig. 3d-f and Supplementary Table S2). The best triple combination (RAP, RIF, ALLAN) doubles mean lifespan, resulting in a median lifespan of up to 44 days and a maximum lifespan of 50 days (Supplementary Table S2). This effect size is comparable to the canonical ageing mutations and is, to the best of our knowledge, the largest lifespan extension ever reported in C. elegans with any adult-onset drug intervention29. By contrast, addition of ALLAN to the non-synergistic combination (RAP+PSORA) did not show any further benefit (Extended data Fig. 3c). Finally, to test whether, despite its high overlap in terms of gene expression changes, addition of MET would results in any further lifespan benefit on dual combinations, we add MET as a third component to one of the synergistic dual combinations RAP+RIF. However this triple combination (RAP+RIF+MET) resulted in toxicity (Extended data Fig. 3b).

supplementary for figure 2. a, RAP+RIF+PSORA did not show further benefit compared to the dual synergistic combinations RAP+RIF and RIF+PSORA. b, RAP+RIF+MET is toxic compared to RAP+RIF. More than 50 worms per condition. c, RAP+PSORA+ALLAN did not show further benefit compared to single drug constituents. d, RAP+RIF+ALLAN and RIF+PSORA+ALLAN resulted in a significant lifespan extension compared to RAP+RIF and RIF+PSORA respectively. log-rank with proper adjustment. e, Mean and f, Maximum lifespan of figure d showed a monotonic increase from single drugs to double and triple combinations. One-way ANOVA, Bonferroni multiple comparison ***P<0.0001, **P<0.001, *P<0.01.

a, all dual and triple synergistic combinations resulted in lifespan extension in daf-16 mutants but effect size does not exceed individual drug effects. b, Of the single drugs only RIF extends lifespan of eat-2 (P<0.05, log-rank with adjustment for multiple comparisons). RIF+PSORA resulted in synergistic lifespan extension in eat-2 mutants (P<0.05, log-rank with adjustment for multiple comparisons). c, None of the synergistic combinations showed synergy in daf-2 mutants. d, RIF alone extends lifespan of daf-7 mutants (P<0.05, log-rank adjustment for multiple comparisons) but combinations fail to result in further synergistic lifespan extension. At least 50 worms per condition.
Mechanisms of drug synergy
Daf-16/FOXO is a transcription factor that plays a key role in lifespan determination in model organisms and likely also in humans30,31. To explore the mode of action of our synergistic drug combinations we determined their dependence on daf-16 in mutant worms deficient in this pathway. Individually, RAP and ALLAN lifespan extension was daf-16-independent while RIF lifespan extension was fully and PSORA was partially daf-16 dependent (Fig. 3a, Extended data Fig. 8). The synergistic combination RAP+RIF comprising one daf-16 dependent (RIF) and one daf-16 independent drug (RAP), and even the RIF+PSORA combination of two daf-16 dependent drugs, still showed synergistic lifespan extension in daf-16 mutants (Fig. 3a, Extended data Fig. 8). This surprising data suggest that synergies can be daf-16 independent, even if individual constituent drugs are daf-16 dependent.
Several of the drugs tested have previously been reported to be CR mimetics17,32-34. We therefore tested each synergistic combination and its constituent compounds in a C. elegans model of CR (eat-2 mutants). We found that only RIF among the single drug treatments further extended lifespan in eat-2 mutants (Fig 3b, Extended data Fig. 8), suggesting that all except RIF may function as CR mimetic in this sense. The synergistic combinations both comprise one CR mimetic with one non-CR mimetic (RAP+RIF and RIF+PSORA). Only RIF+PSORA resulted in further lifespan extension and addition of ALLAN, a second CR mimetic, did not result in further benefits.
To further explore the mechanism of drug synergy we determine the transcriptome profile for each synergistic combination and selected non-synergistic drug combinations. Transcriptomic analysis in worms treated with single, double and triple drug exposure for the synergistic and non-synergistic combinations revealed that the transcriptome of CR mimetic single drugs is clustered together with eat-2, whereas the non-CR mimetic RIF is different from eat-2 and from all other single drugs (Fig. 2h). Interestingly, the transcriptomes of synergistic combinations are clustered together and the pattern is different from those of their constituent single drugs and from that of eat-2 mutants (Fig. 2h, Extended data Fig. 4c,d). Moreover, we found that only on pathway, transforming growth factor beta (TGFβ), was commonly enriched in all synergistic dual and triple combinations (Fig. 4a, Extended data Fig. 6c, 7a, Supplementary Table S4) relative to untreated control. To identify pathways that were further enriched by the drug combinations relative to their constituent single drugs, we used the single drug transcriptome as background control (see methods) for pathway analysis. Using this approach, we found that all synergistic combinations, again, affected TGFβ amongst the pathways differentially impacted relative to their parent drugs while none of the non-synergistic combinations did so (Supplementary Table S5). Because some drug combinations resulted in further lifespan extension in eat-2 mutants relative to their constituent drugs, we tested whether TGFβ was enriched in the transcriptomes of N2 animals treated with synergistic drug combinations relative to untreated eat-2 worms. Again, TGFβ was the only pathway enriched in all synergistic combinations (Extended data Fig. 7b Supplementary Table S6). Overall, we found that the TGFβ signaling pathway was commonly and exclusively enriched in synergistic combinations (Fig. 4b, Extended data Fig. 7a,c, Supplementary Table S7).

a, 2D PCA of PSORA, RIF and their combination showed that the combination transcriptome profile is different from single drugs. Because the single drugs and the combinations were sequenced in two different lanes, we used the N2 untreated controls of both lanes. b, as in a for RAP and RIF. c, similar to the 3D PCA, RAP, RIF and PSORA were well separated with 2D PCA. All single drugs were separated from eat-2. d, all synergistic combinations are well separated with N2 control and eat-2, whereas they are not separated from each other. This could show that they may have the same mechanism of synergistic lifespan extension.


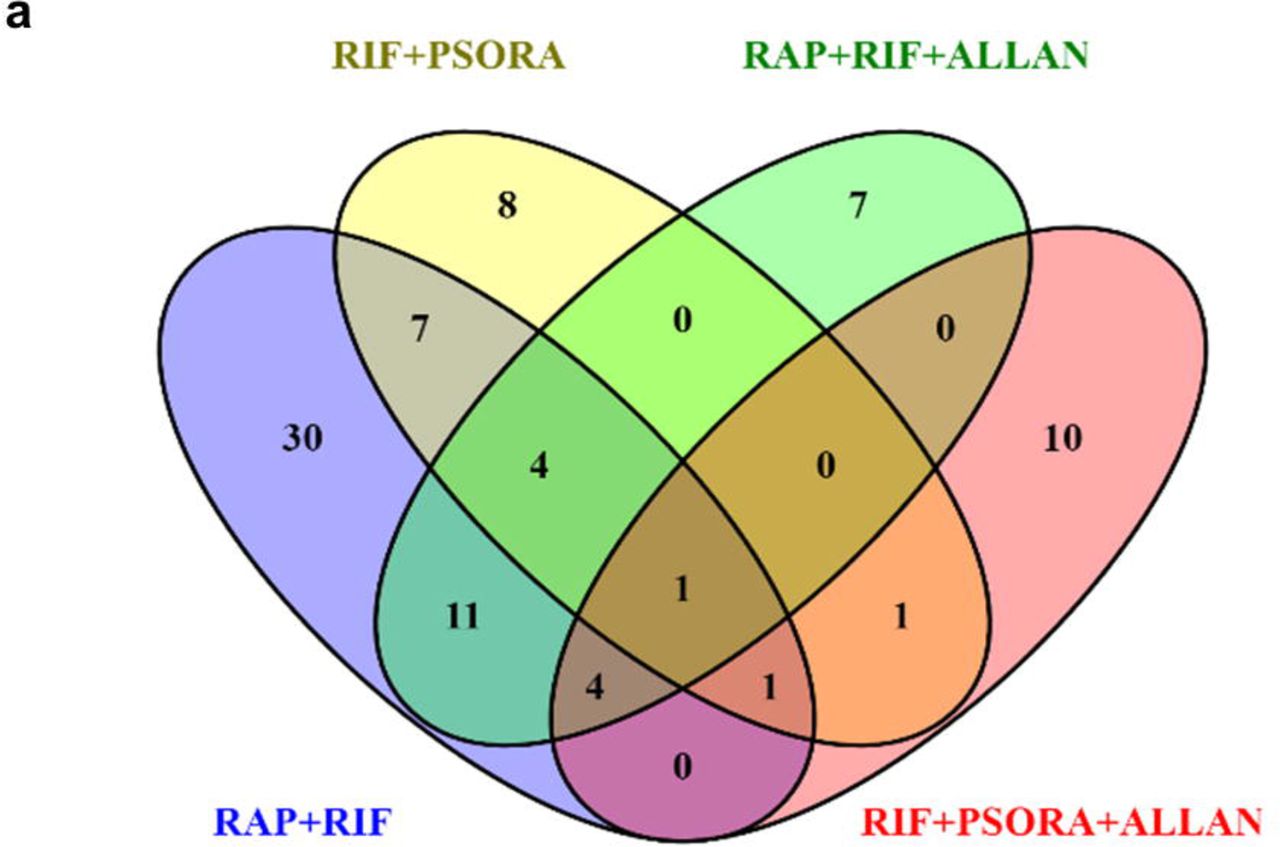
a, DGE and GO term overlap among synergistic combinations. Purple line links identical genes whereas blue line links different gene that grouped into similar GO terms. b, Venn diagram of pathways enriched in synergistic combinations using eat-2 transcriptome as a background. Only TGF-beta was commonly enriched in all synergistic combinations.

a, Top five pathways enriched in the down regulated genes of the four synergistic drug combinations. All synergistic combinations consistently enrich TGFβ. b, Pathways enriched in N2 animals treated with synergistic drug combinations relative to untreated eat-2 worms. All synergistic combinations consistently enrich TGFβ. c, fold enrichment of TGFβ in all single drugs and synergistic combinations. From single drugs, only RIF enriched TGFβ. All the four synergistic combinations resulted in a significant enrichment of TGFβ compared to RIF. d, protein-protein interaction drawn by STRING72 showed that TGFβ/daf-7 has an interaction with daf-2 and daf-16 as well as the lipid metabolism regulators daf-9 and daf-12. Thickness of the edge indicates strength of data support for the specific interaction. e, PC’s pooled based on their degree of unsaturation, an increase shown in the MUFA containing species normalized to control, 2500 worms per condition (Mean ± SD) *P<0.01, **P<0.001, ***P<0.000.





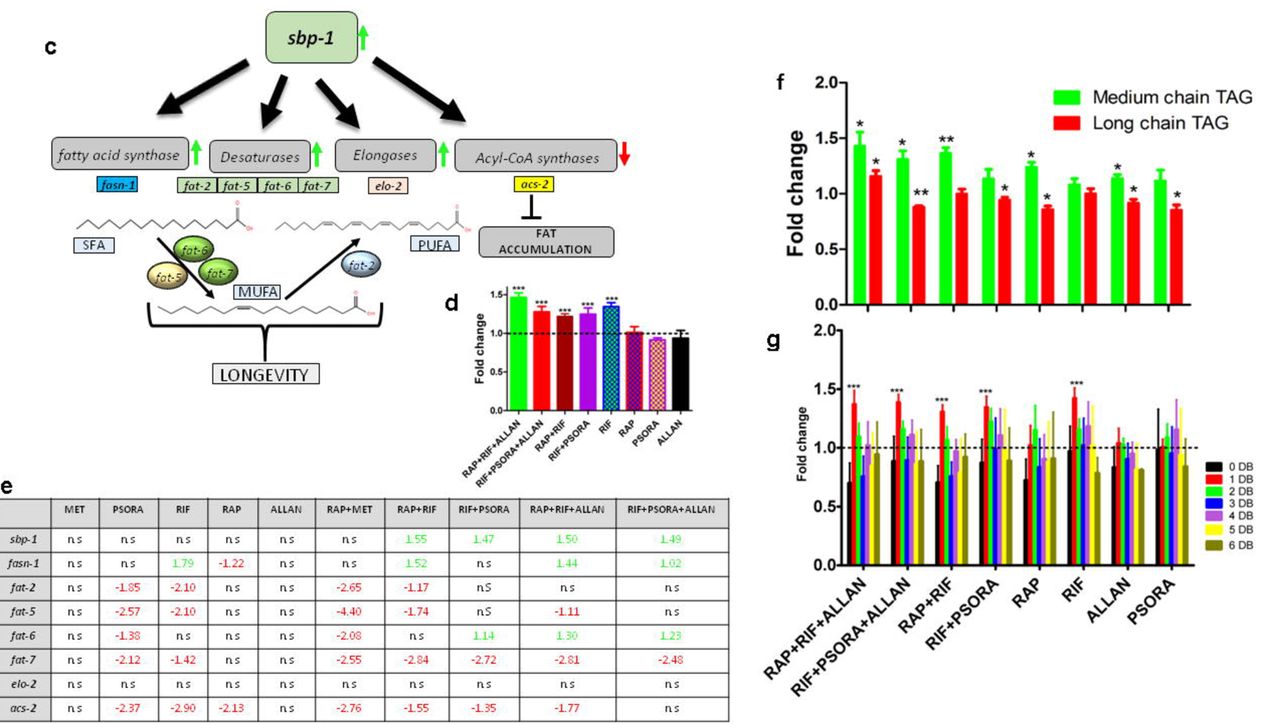
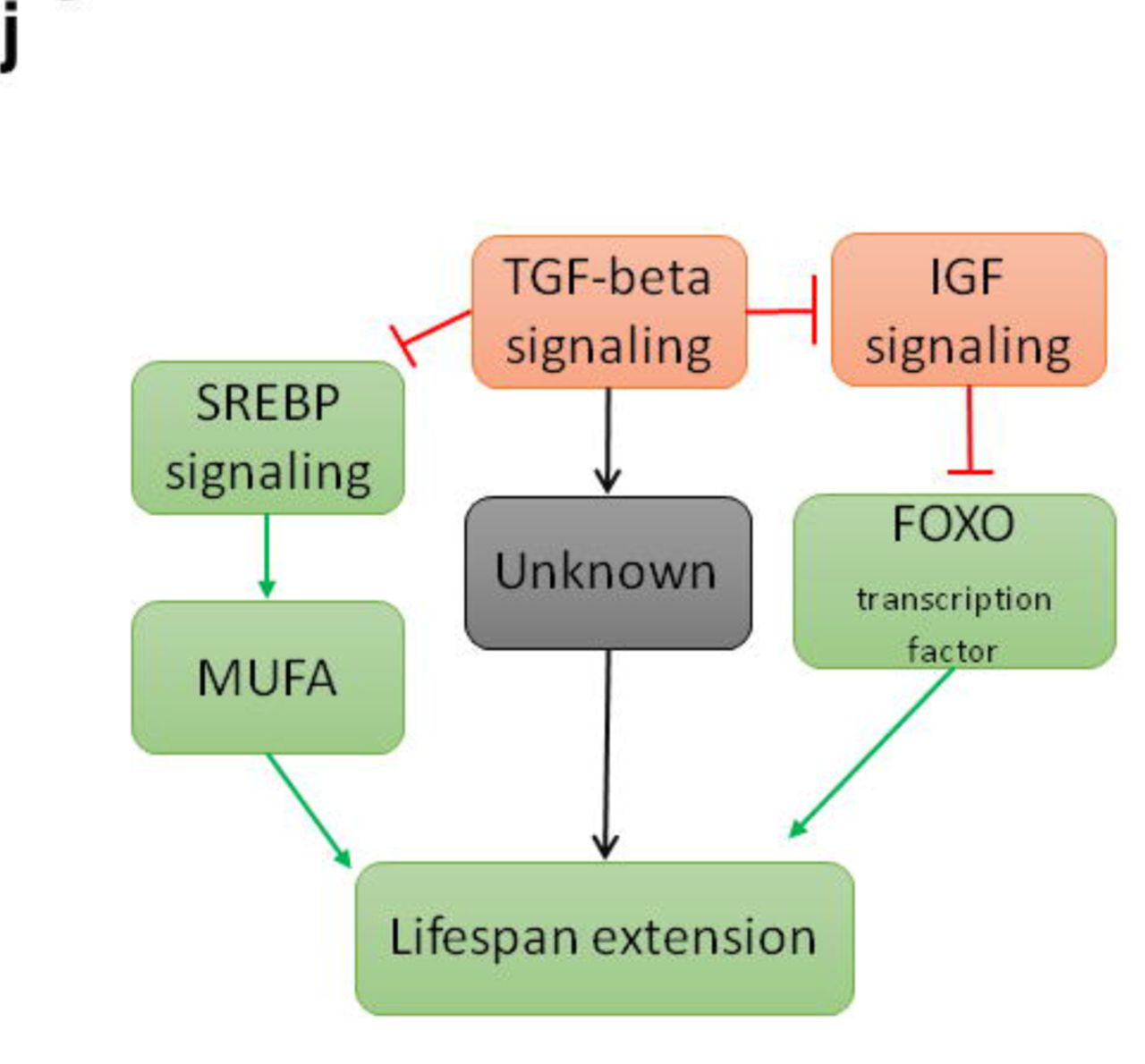
a, Venn diagram for pathways enriched by synergistic drug combinations. Only is commonly enriched in all synergistic combinations. b, Heatmap of pathways enriched by synergistic drug combinations compared to their constituent single drugs. Only is commonly enriched in all synergistic combinations. c, Mechanism of MUFA containing lipid accumulation and lifespan extension. d, Total triacylglycerol content normalized to the control, three biological replicates, 2500 worms per condition, (Mean ± SD, one-way ANOVA) e, Fat metabolism-related gene expression profile, log fold change, P value < 0.05. f, TAGs categorized as those containing medium chained and long chained acyl chains, normalized to control, three biological replicates, 2500 worms per condition (Mean ± SD, one-way ANOVA). g, Phosphatidylcholine species abundance based on double bond, three biological replicates, 2500 worms per condition (Mean ± SD, one-way ANOVA). h, Total sphingomyelin content normalized to the control, three biological replicates, 2500 worms per condition, (Mean ± SD, one-way ANOVA). i, Double bond index, per oxidation index and MUFA/PUFA ratios calculated for phosphatidylcholine species for all conditions, MUFA/PUFA is normalized to the control. Three biological replicates, 2500 worms per condition (Mean ± SD), j. Hypothesized mechanism of lifespan extension by drug synergies. Red–inhibition, green–activation, black–unknown. *P<0.01, **P<0.001, ***P<0.0001.
TGFβ is required for drug synergy
The specific and consistent enrichment of TGFβ suggests that TGFβ may play a role in mediating the observed synergistic lifespan extension. Previously it has been shown that TGFβ (daf-7) mutation extends lifespan via insulin signaling and that both daf-2 and daf-7 regulate the transcription of daf-16 dependent genes35 (Extended data Fig. 7d). To test the role of the IGF pathway in this effect and to determine if the lifespan of already long-lived daf-2 mutants could be further extended using our drug combinations, we first determined lifespan of daf-2 mutants for each combination. Only the RAP-based combinations, RAP+RIF and RAP+RIF+ALLAN, showed lifespan extension in daf-2 mutants. However, the effect size was similar to that of RAP alone and none of the combinations resulted in further synergy (Fig. 3c, Extended data Fig. 8, Supplementary Table S2). PSORA treatment alone extended the lifespan of daf-2 mutants, but none of the PSORA based combinations, RIF+PSORA and RIF+PSORA+ALLAN, extended lifespan in daf-2 mutants (Fig. 3c and Extended data Fig. 8). To test our hypothesis that TGFβ was involved in mediating drug synergies, we next tested the efficacy of all synergistic combinations and their components in daf-7 mutants. None of the combinations show synergistic effects in the daf-7 mutants, suggesting that synergy requires daf-7, even though some of the drugs singularly still extend lifespan (Fig. 3d, Extended data Fig. 8).

a, Dual combinations RAP+RIF and RIF+PSORA showed synergy in daf-16 where as the triple combinations RAP+RIF+ALLAN and RIF+PSORA+ALLAN did not result in significant difference from their respective synergistic dual combinations. b, All synergistic combinations did not show synergy in daf-2 mutant worms. c, Only RIF+PSORA showed synergy in eat-2 mutants. d, The triple combinations showed further lifespan extension compared to single drugs but none of the four combinations show synergy in daf-7 mutants. Supplementary for figure 4. At least 50 worms per condition. One-way ANOVA, Bonferroni multiple comparison ***P<0.0001, **P<0.001, *P<0.01.
Transcriptome, Lipidome and Drug synergy
Long-lived mutants such as age-1 and daf-2 have previously been shown to exhibit metabolic perturbations resulting in increased production and storage of fats36. Furthermore, TGFβ/daf-7 regulates triacylglycerol (TAG) metabolism37 and daf-7 mutants are known to store more fats36. These links in conjunction with our transcriptomics data led us to investigate whether treatment with synergistic drug combinations resulted in modifications in lipid profiles consistent with TGF inhibition. (Supplementary Table S3). First we explored the transcriptomics data for changes that might affect lipid composition and found that synergistic drug combinations indeed resulted in a significant up regulation of the C. elegans SREBP-1c homolog - sbp-1, a master transcription factor controlling several lipogenic genes, and genes coding for “desaturases” responsible for MUFA synthesis38-40 (Fig. 4c,e). We then employed an MS-based lipidomics assay to determine changes in lipid compositions that might result from these drug treatments. Worms that were exposed to the synergistic drug combinations have a significant rise in TAG reserves (Fig. 4d), with increased abundances in those TAG species that contained the medium chained saturated fatty acids (Fig. 4f). In addition, we also observed a marked increase in the MUFA:PUFA ratios (Fig 4g, Extended data Fig. 7e). Because of these changes, there was a significant decline in lipid peroxidation index (PI) (Fig. 4i). A low PI, indicates that lipids contain fewer carbon-carbon double bonds, making them less susceptible to peroxidation and this has been previously associated with increased lifespan, probably related to lower susceptibility to lipid peroxidation41,42,43. A reduction in the susceptibility to lipid peroxidation suggests better resistance to oxidative stress44,45 and worms treated with synergistic drug combinations indeed showed such an increase in resistance (Fig. 5f). We also observed an increase in total sphingomyelin (Fig. 4h); an event that has previously shown to elicit autophagy-dependent lifespan extension in the nematodes46.

a-c, RIF+PSORA+ALLAN and RAP+RIF+ALLAN shows extension of reproductive span but no effect on total fertility, n=10 worms per condition. d, health span, n>150 worms per condition, P<0.0001, one-way ANOVA. e, resistance to heat stress, n=50 worms per condition. f, resistance to paraquat stress, n=50 worms per condition. g, distance travelled n=50 worms per condition, one-way ANOVA. h, development measured by size of worms n=10 worms per condition. i, basal metabolic rate, n=80 worms per condition, one-way ANOVA, Bonferroni multiple comparison. *P<0.01, **P<0.001, ***P<0.0001
Drug synergy improves health span and delays ageing
Suppression of energy metabolism and inhibition of the electron transport chain (ETC) can extend lifespan in C. elegans47,48. In our experiments, however, drug treatments did not have inhibitory effects on basal metabolic rate, instead several of this drugs and drug combinations increased basal respiration (Fig. 5i), excluding ETC inhibition as explanation for the observed lifespan effects. The most common tradeoffs related with lifespan extension are developmental delay, slower growth rate and reduced fecundity49,50,51. We found that in worms treated with either of the synergistic drug combinations, there is evidence for a delay in peak egg laying and an extension of reproductive span but no change in total fertility. (Fig. 5a-c, Extended data Fig. 9a).

a, Drug synergies extended reproductive period. 10 worms per condition. b, Heat shock stress, RIF+PSORA+ALLAN improves heat shock stress resistance both at young, day 4, worms and older, day 15, worms c, Health span scoring at different age. At all ages from young to old drug synergy treated worms have better health span than their respective age matched controls. ***P<0.0001, **P<0.001, *P<0.01.
Next, we assessed several parameters of fitness, stress resistance and health: speed of spontaneous movement, tolerance to heat shock and oxidative stress resistance and a motility based health span score based on the scoring scheme of Herndon et al52. Control animals spent about 50% of their lifespan in the optimal (best) health category while nematodes exposed to the RIF+PSORA+ALLAN and RAP+RIF+ALLAN spent 57% and 63%, respectively, of their already extended lifespan in optimal health (Fig. 5d, Extended data 9c). This means that their health span was significantly extended both in absolute and relative terms. Indeed, more than half of the treated animals were still in optimal health even after the last control animal had died (Fig. 5d, Extended data Fig. 9c). Surprisingly, animals treated with RAP+RIF+ALLAN and RIF+PSORA+ALLAN also performed significantly better in the spontaneous movement assay than control animals at most ages. In fact, old treated animals (day 18 of age) showed performance in this assay that was indistinguishable from young (day 7) controls (Fig. 5g). Moreover, treated animals experienced improvements in resistance to heat and oxidative stress (paraquat) assays, (Figure 5e,f, Extended data Fig. 9b). These observations indicate that animals treated with synergistic drug combinations not only enjoyed greater lifespan with limited detected tradeoffs, but also improved health span, increased stress resistance, and even increased physical performance. Such observations raise the important question if these lifespan benefits are due to an actual delay in biological ageing rate. The rate of ageing can be expressed as mortality rate doubling time (MRDT). We determined MRDT of animals exposed to synergistic drug combinations using Survcurv53, and found MRDT to be significantly longer in RIF+PSORA+ALLAN treated animals compared to controls (MRDT of control = 3 days and RIF+PSORA+ALLAN = 3.7 days, P value < 0.0001). The initial mortality rate was significantly lower for both synergistic combinations ((IMR of control = 2.7e-3, IMR of RIF+PSORA+ALLAN = 8.5e-4, RAP+RIF+ALLAN = 9.3e-4, P value < 0.001). This means that the RIF+PSORA+ALLAN drug combinations extend lifespan by making worms age more slowly while at the same time making them move faster/increasing spontaneous activity and increasing their resistance to stress when young. (Extended data Fig. 10).

a, Both the dual and triple synergistic combinations slow the rate of mortality of wild type N2 worms. b, The triple combinations RAP+RIF+ALLAN and RIF+PSORA+ALLAN slows mortality rate in Drosophila Melanogaster. c, RAP+RIF+ALLAN and d, RIF+PSORA+ALLAN delays incidence rate of death of worms.
Drug synergy is conserved in Drosophila
One of the key questions for ageing studies is whether lifespan effects are conserved across species. We therefore tested whether the synergistic lifespan extensions we found in C. elegans are conserved in Drosophila melanogaster. RAP has previously been shown to extend lifespan of fruit flies54 while MET has been shown to be ineffective55. We systematically tested all single drugs and all synergistic combinations in male fruit flies. First, we showed that RAP and ALLAN extend mean lifespan whereas RIF and PSORA extend only the maximum lifespan of flies (Fig. 6 Supplementary Table S2). However, the RAP based combinations RAP+RIF and RAP+RIF+ALLAN showed conserved synergistic lifespan extension in flies (Fig. 6a). The PSORA based combinations; RIF+PSORA and RIF+PSORA+ALLAN did not show further mean lifespan extension (Fig. 6b). However, RIF+PSORA showed synergistic maximum lifespan extension compared to single drugs (Fig. 6c). Comparison of the Gompertz parameters of the mortality trajectories (Extended data Fig. 10b) showed that the baseline mortality rate was not significantly different in drug combination treated flies (IMR difference of RAP+RIF+ALLAN = 2.9e-1, RIF+PSORA+ALLAN = 4.8e-1, NS) compared to untreated control. However, the rate of increase of mortality with age is significantly lower in drug combination treatments (Gompertz different b of RAP+RIF+ALLAN = 2.20e-4, P-value < 0.001, Gompertz different b of RIF+PSORA+ALLAN = 1.15e-2, P-value <0.05) relative to the control. It should be noted that cohort size of flies was smaller than for worms and that we did not re-optimize drugs for the fly experiments. However, these data confirms that some of the detected synergies appear to be evolutionary conserved.

a,b,c RAP extends both mean and maximum lifespan. RAP+RIF and RAP+RIF+ALLAN resulted in synergistic mean and maximum lifespan extension. RIF and PSORA did not extend mean lifespan of flies. Their combination also did not extend mean lifespan. However, RIF and PSORA extends maximum lifespan significantly. RIF+PSORA and RIF+PSORA+ALLAN resulted in further maximum lifespan extension, close to additive (P value < 0.001) but not mean lifespan. n = 80 flies per condition. One-way ANOVA, Bonferroni multiple comparison for mean and maximum lifespan and Log-rank test for survival analysis. *P<0.01, **P<0.001, ***P<0.0001
Discussion
By targeting multiple overlapping ageing and longevity-related pathways we succeeded in designing synergistic pharmacological interventions that, even when only applied to animals from adult age, more than doubled healthy lifespan in C. elegans. This effect size is comparable to that of the classical ageing mutations and, to the best of our knowledge, is the largest reported for any adult-onset pharmacological intervention13. However, unlike in ageing mutations we detect very limited detrimental evolutionary or fitness tradeoffs associated with this lifespan extension by drug synergies 19. In particular, we did not detect any decrease in oxygen consumption or total fecundity. The latter result is consistent with a previous study showing benefits of an adult-onset intervention using resveratrol without reduced fecundity56. These data suggest that there is significant post-developmental plasticity of lifespan that can be exploited using adult-onset pharmacological interventions. In fact, treated animals outperform age-matched controls in some performance assays and old treated animals have the physical appearance and performance comparable to much younger controls (Fig. 5g). Mortality analysis suggests that one synergistic drug treatment slows basic biological ageing rate by ~20% (Extended data Fig. 10). To test for evolutionary conservation, we determined lifespan benefits in the fruit fly Drosophila melanogaster, confirming qualitative conservation for two of the four synergistic drug combinations. Nematodes are evolutionarily more distant from fruit flies than they are from mammals57, making mechanisms and pathways conserved between flies and nematodes ancient, and tracing this synergy back to a common ancestor of all three clades.
To explore the mechanism of these synergistic lifespan benefits, we carried out epistasis experiments and transcriptomic analysis, finding that daf-2/daf-16 (IGF/FOXO) as well as the TGFβ pathway (daf-7) are involved in synergistic lifespan extension (Fig. 4j). The connection between IGF/FOXO and TGF is consistent with previous results where daf-7 regulates lifespan via insulin signaling35, neuronal TGFβ links food availability and longevity58, and daf-7 regulates metabolism in C. elegans and controls fat accumulation36. Several previous studies have demonstrated that lifespan extending mutations in C. elegans result in alterations in lipid metabolism causing an increase in the MUFA and reduction in PUFA levels59,60. In agreement with these reports, our drug synergies resulted in an up regulation of sbp-1, a master transcription factor controlling several genes related to lipid metabolism and MUFA synthesis38-40 (Fig. 4c,e, Extended data Fig. 5). MS-based lipidomics analysis of nematodes exposed to synergistic drug combinations revealed major changes consistent with activation of sbp-1, including accumulation of TAGs and increases in the MUFA:PUFA ratios (both in PC and PE classes). Previously, it has been shown that an increased TAG abundance prolongs lifespan in budding yeast61 and C. elegans36,38. Furthermore, upon closer inspection of the fatty acyl chains bound to the TAG species, we found a significant increase in those that contained medium chained saturated fatty acyls (MCFA) (14:0, 16:0), (Fig. 4f). The findings from lipidomics therefore are consistent with the observed induction of the sbp-1 and its target lipogenic genes (Figure 4c). Increased abundance of such medium chained saturated fatty acyls has also been reported to be positively correlated with longevity in C. elegans previously59. Similarly, high abundance of MUFAs has been shown to be associated with extended lifespan in nematodes62. In humans, high ratios of MUFA:PUFA have also been found in erythrocyte membranes of children of nonagenarians63,64. There has been much interest in the potential for MET and possibly RAP to delay age-dependent decline and disease in humans. Given this translational interest, the limited benefit of combining MET with RAP in both C. elegans and mice is somewhat disappointing. However, as our results illustrate, lifespan is determined through complex and interactive biochemical and gene regulatory networks. Intervening simultaneously at multiple points of these networks can result in significant and sometimes surprising benefits28. Our proof-of-principle study suggests that beneficial synergistic and additive interactions affecting key longevity pathways are unexpectedly common and evolutionarily conserved. While, to date, ours is the largest systematic transcriptional screen of drug synergy interactions between compounds targeting ageing pathways, it is important to note that this dataset is too small to derive definitive rules by which to detect or predict synergistic interactions based on the transcriptional profiles of single drugs or even drug combinations. Moreover, there is growing interest in repurposing drugs by identifying compounds that may target known or suggested ageing pathway based on the often-extensive data known about such compounds, including their three-dimensional structure bound to biological ligands, their pharmacokinetics, physical properties and known biological targets and effects65. Our finding support the feasibility of targeting multiple conserved ageing pathways using existing drugs to slow down biological ageing rate, an approach that, if translatable to humans, would result in dramatic medical and economic benefits66.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}