

Abstract
One Sentence Summary Single-molecule imaging reveals that eukaryotic condensin is a highly processive DNA-translocating motor complex.
Abstract Condensin plays crucial roles in chromosome organization and compaction, but the mechanistic basis for its functions remains obscure. Here, we use single-molecule imaging to demonstrate that Saccharomyces cerevisiae condensin is a molecular motor capable of ATP hydrolysis-dependent translocation along double-stranded DNA. Condensin’s translocation activity is rapid and highly processive, with individual complexes traveling an average distance of ≥10 kilobases at a velocity of ∼60 base pairs per second. Our results suggest that condensin may take steps comparable in length to its ∼50-nanometer coiled-coil subunits, suggestive of a translocation mechanism that is distinct from any reported DNA motor protein. The finding that condensin is a mechanochemical motor has important implications for understanding the mechanisms of chromosome organization and condensation.
Structural maintenance of chromosomes (SMC) complexes are the major organizers of chromosomes in all living organisms (1, 2). These protein complexes play essential roles in sister chromatid cohesion, chromosome condensation and segregation, DNA replication, DNA damage repair, and gene expression. A distinguishing feature of SMC complexes is their large ring-like architecture, the circumference of which is made up of two SMC protein coiled-coil proteins and a single kleisin subunit (Fig.1A) (1-4). The ∼50-nm long antiparallel coiled-coils are connected at one end by a stable dimerization interface, referred to as the hinge domain, and at the other end by globular ATP-binding cassette (ABC) family ATPase domains (5). The ATPase domains are bound by a protein of the kleisin family, along with additional accessory subunits, which vary for different types of SMC complexes (Fig. 1A). The relationship between SMC structures and their functions in chromosome organization is not completely understood (6), but many models envision that the coiled-coil domains allow the complexes to topologically embrace DNA (1-4). Given the general resemblance to myosin and kinesin, some early models postulated that SMC proteins might be mechanochemical motors (7-10).

A) Schematic of the S. cerevisiae condensin complex. The Brn1 kleisin subunits connects the ATPase head domains of the Smc2–Smc4 heterodimer and recruits the HEAT-repeat subunits Ycs4 and Ycg1. The cartoon highlights the position of the HA3-tag used for labeling. (B) Conceptual schematic of loop extrusion models. (C) Wild-type and ATPase-deficient Smc2(Q147L)–Smc4(Q302L) condensin complexes analyzed by SDS PAGE and Coomassie staining. (D) Electron micrographs of wild-type condensin holocomplexes rotary shadowed with Pt/C. Scale bar: 100 nm. (E) Electrophoretic mobility shift assays with 6-FAM labelled 45-bp dsDNA substrate (100 nM) and the indicated protein concentrations. (F) ATP hydrolysis by wild-type and ATPase mutant condensin complexes (0.5 μM) upon addition of increasing concentrations of a 6.4-kb linear DNA at saturated ATP concentrations (5 mM). The plot shows mean ± standard deviation of n = 3 (wild-type) or 2 (ATPase mutant) independent experiments. (G) Michaelis-Menten kinetics for the rate of ATP hydrolysis by wild-type condensin complexes (0.5 μM) at increasing ATP concentrations in the presence of 240 nM 6.4-kb linear DNA. The plot shows mean ± S.D. of N = 3 independent experiments. The fit corresponds to a Km of 0.4 ± 0.07 mM for ATP and kcat of 2.0 ± 0.1 s-1 per molecule of condensin (mean ± standard error).
SMC complexes are thought to regulate genome architecture by physically linking distal chromosomal loci, but how these bridging interactions might be formed remains unknown (1, 2, 11). An early model suggested that many three-dimensional (3D) features of eukaryotic chromosomes might be explained by DNA loop extrusion (Fig. 1B) (12), and recent polymer dynamics simulations have shown that loop extrusion can recapitulate the formation of topologically associating domains (TADs), chromatin compaction, and sister chromatid segregation (13-17). This loop extrusion model assumes a central role for SMC complexes in actively creating the DNA loops (11, 12). Similarly, it has been proposed that prokaryotic SMC proteins may structure bacterial chromosomes through an active loop extrusion mechanism (18). Yet, the loop extrusion model remains hypothetical, in large part because the motor activity that is necessary for driving loop extrusion could not be identified (11). Indeed, the absence of an identifiable motor activity in SMC complexes instead has lent support to alternative models in which DNA loops are not actively extruded, but instead are captured and stabilized by stochastic pairwise SMC binding interactions to bridge distal loci (19).
To help distinguish between possible mechanisms of SMC protein-mediated chromosomal organization, we examined the DNA-binding properties of condensin (20). We overexpressed the five subunits of the condensin complex in budding yeast and purified the complex to homogeneity (Fig. 1C and fig. S1). Electron microscopy images confirmed that the complexes were monodisperse (Fig. 1D). As previously described for electron micrographs of immunopurified Xenopus laevis or human condensin (21), we observed electron density that presumably corresponds to the two HEAT-repeat subunits in close vicinity of the Smc2–Smc4 ATPase head domains. We confirmed that the S. cerevisiae condensin holocomplex binds double-stranded (ds) DNA and hydrolyzes ATP in vitro (Fig. 1E and F). Addition of dsDNA stimulated the condensin ATPase activity ∼3-fold, which is consistent with previous measurements with X. laevis condensin I complexes (22), and revealed Km and kcat values of 0.4 ± 0.07 mM and 2.0 ± 0.1 s-1, respectively, for ATP hydrolysis in the presence of linear dsDNA (Fig. 1G). An ATPase-deficient version of condensin with mutations in the γ-phosphate switch loops (Q-loops) of Smc2 and Smc4 still bound DNA (Fig. 1E), but exhibited no ATP hydrolysis activity (Fig. 1F).
We then used total internal reflection fluorescence microscopy (TIRFM) to visualize condensin binding to DNA curtains at the single-molecule level (23). Initial experiments using single-tethered DNA curtains demonstrated that condensin could promote extensive ATP hydrolysis-dependent DNA compaction, which was reversible by increasing the salt concentration to 0.5 M NaCl (Fig. S2). We next asked whether we could directly visualize the binding of single fluorescently-tagged condensin holocomplexes to double-tethered DNA substrates. We fluorescently labeled condensin with quantum dots (Qdots) conjugated to antibodies against the HA3-tag fused to the Brn1 kleisin subunit (Fig. 1A). Electrophoretic mobility shift assays confirmed that condensin was quantitatively labeled (Fig. S3A). Importantly, binding to the Qdots inhibited neither condensin’s ATP hydrolysis activity nor its ability to alter DNA topology (Fig. S3B and C). We prepared double-tethered curtains by attaching the DNA substrates (∼48.5-kb λ-DNA) to a supported lipid bilayer through a biotin-streptavidin linkage, and aligned one end of the DNA molecules at nanofabricated chromium (Cr) barriers and anchored the other end of the DNA to Cr pedestals located 12 μm downstream (Fig. 2A) (23).

(A) Schematic of the double-tethered DNA curtain assay. (B) Still images showing Qdot-tagged condensin (magenta) bound to YoYo1-stained DNA (green). (C) Kymograph showing examples of Qdot-tagged condensin translocating on a single DNA molecule (unlabeled); the initial condensin binding sites, dissociation positions, and collisions with the barriers/pedestals are highlighted with color-coded arrowheads. (D) Kymograph showing Qdot-tagged ATPase-deficient mutant Smc2(Q147L)– Smc4(Q302L) condensin undergoing 1D diffusion on DNA (unlabeled). (E) Initial binding sites and (F) dissociation site distributions of condensin superimposed on the AT content of the λ-DNA substrate. All reactions contained 4 mM ATP. Error bars in (E) and (F) represent standard deviations calculated by boot strap analysis.
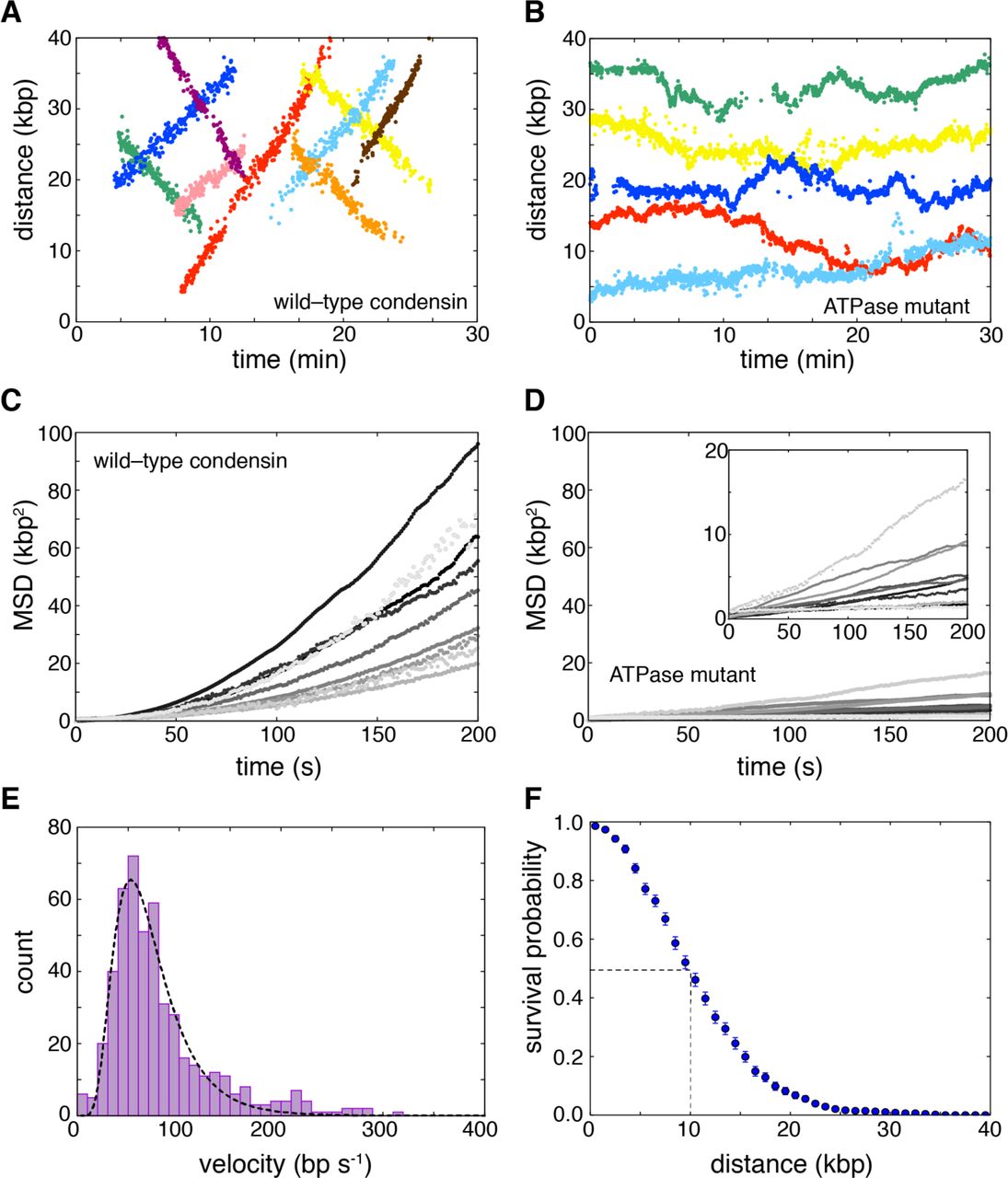
Using double-tethered curtains, we were able to detect binding of condensin complexes to individual DNA molecules (Fig. 2B). Kymographs revealed that, remarkably, ∼85% of all bound condensin complexes (N = 671) underwent linear motion along the DNA (Fig. 2C). The up/down direction of movement was observed to be random, but once a complex started translocation, it did generally proceed unilaterally without a reversal of direction (reversals were observed occasionally, in 6% of the traces). Condensin has not been previously shown to act as a molecular motor, but the observed movement is fully consistent with expectations for an ATP-dependent DNA-translocating motor protein. The ATPase-deficient Q-loop mutant of condensin only exhibited motion consistent with random 1-dimensional (1D) diffusion (Fig. 2D). Wild-type condensin in the presence of the non-hydrolyzable ATP analog ATPγS similarly only displayed 1D-diffusion (Fig. S4A). Analysis of the initial binding positions for wild-type condensin revealed a preferential binding to A/T-rich regions (Pearson’s r = 0.66, P = 5×10−6; Fig. 2E), similar to values reported for Schizosaccharomyces pombe cohesin (24). In contrast, the condensin dissociation positions were not correlated with A/T content (Pearson’s r = –0.05, P = 0.77), nor were there any other preferred regions for dissociation within the DNA, with the exception of the Cr barriers and pedestals (Fig. 2F). These findings are consistent with a model where condensin loads at A/T-rich sequences and then translocates away. Interestingly, previous single-molecule experiments demonstrated rapid 1D diffusion of cohesin on DNA, but found no evidence for ATP-dependent translocation, suggesting that there may be a difference between how the two SMC complexes process DNA (24, 25).
We used particle tracking to quantitatively analyze the movement of condensin on DNA (Fig. 3A and B, and fig. 4SB). Wild-type condensin did not travel in a preferred direction: 52% (N=255/491) went one direction, and 48% went the opposite direction (N=236/491). The condensin ATPase mutant did not exhibit any evidence of unidirectional translocation. Mean squared displacement (MSD) plots generated from condensin tracking data revealed increasing slopes (Fig. 3C), which is only consistent with directed motion (26). In contrast, MSD plots were linear for the ATPase-deficient condensin mutant (Fig. 3D) and for wild-type condensin in the presence of ATPγS (Fig. S4C). Linear MSD plots were characteristic of random diffusive motion (26), yielding diffusion coefficients (D1,obs) of (1.7 ± 1.4)×10−3 and (0.8 ± 1.0)×10−3 μm2 s-1 (mean±S.D.) for ATPase-deficient condensin and wild-type condensin plus ATPγS, respectively.

(A) Minimal mechanistic framework necessary for coupling ATP-dependent translocation to the extrusion of a cis DNA loop. In this generic model, a motor domain (green) must move away from a second DNA-binding domain (blue), and this secondary domain can either remain stationary (as depicted) or it may also act as a motor domain and move in the opposite direction (not depicted). (B) Detection of Cis loop extrusion is not possible when the DNA is held in a fixed configuration as in the double-tethered curtains that allows for direct detection of condensin motor activity in the absence of condensation (top panel). The middle and bottom panels show a schematic of an assay to mimic cis DNA loop extrusion by providing a second λ-DNA substrate in trans. (C) Examples of kymographs showing translocation of a second λ-DNA substrate (stained with YoYo1) provided in trans in the presence of unlabeled condensin. The presence of the trans DNA substrate is revealed as regions of locally high YoYo1 signal intensity, as highlighted by arrowheads. The regions of higher signal intensity are not detected when the trans DNA is omitted from the reaction. (D) Velocity distribution histogram and (E) survival probability plot for condensin bound to the trans DNA substrate. The dashed line in (E) highlights the translocation distance corresponding to dissociation of one half of the bound condesin complexes. Error bars represent standard deviations calculated by boot strap analysis. Cartoons of generalized models for condensin motor activity through a (F) “scrunching” or (G) “walking” mechanisms, both of which can be based upon ATP hydrolysis-dependent changes in the geometry of the SMC coiled-coil domains. Models are discussed in the text.

A) Examples of tracked translocation trajectories for Qdot-tagged wild-type condensin and (B) the ATPase-deficient Smc2(Q147L)–Smc4(Q302L) condensin mutant. Mean squared displacement (MSD) plots for (C) wild-type condensin and (D) the ATPase deficient mutant obtained from the tracked trajectories. (E) Velocity distributions for condensin translocation activity; the dashed line is a log-normal fit to the translocation rate data. (F) Processivity measurements of condensin motor activity. The dashed line highlights the translocation distance corresponding to dissociation of one half of the bound condesin complexes. Error bars represent standard deviations calculated by boot strap analysis.
We used the tracking data to determine the velocity and processivity for wild-type condensin. A plot of the velocity distributions for data collected in the presence of saturated concentrations of ATP (4 mM; Fig. 1G) was well described by a log-normal distribution, revealing a mean apparent translocation velocity of 63 ± 36 bp s-1 (16 ± 9 nm s-1) (mean±S.D.) (Fig. 3E). Interestingly, upon initial binding, condensin paused for a brief period (τpause=13.3 ± 1.5 s) before beginning to move along the DNA, which suggests the existence of a rate-limiting step prior to becoming active for translocation (Fig. 2C and fig. S5). Each translocating condensin complex remained bound to the DNA for an average total time of 4.7 ± 0.2 min and traveled on average 10.3 ± 0.4 kb (2.6 ± 0.1 μm) before dissociating (Figs. 3F and fig. S6A). This value provides merely a lower limit of the processivity of condensin, because a significant fraction (42%) of the complexes traveled all the way to the ends of the 48.5-kb λ-DNA, where they collided with the Cr barriers or pedestals (see, for example, Fig. 2C). There was no correlation between translocation velocity and processivity at a given ATP concentration (Pearson’s r = 0.035, P = 0.43 at 4 mM ATP) (Fig. S6B). However, velocity and processivity both varied with ATP concentrations. Michaelis-Menten analysis revealed a vmax of 62 ± 2 bp s-1 and a Km of 0.2 ± 0.04 mM ATP (Fig. S7A and B). The initial pause time (τpause) also varied with ATP concentration, from a mean value of 3.9 ± 0.8 min at 50 μM ATP to 13.3 ± 1.5 s at 4 mM ATP, suggesting that this delay reflects a transition from a translocation-inactive to translocation-active state that is dependent upon ATP binding, ATP hydrolysis, or both (Fig. S7C).
Our finding that condensin is an ATP hydrolysis-dependent molecular motor lends support to models invoking SMC protein-mediated loop extrusion as a means for 3D genome organization. An important prediction of the loop extrusion model is that condensin must simultaneously interact with two distal regions of the same chromosome, and at least one (or possibly both) of the interaction sites must translocate away from the other site, allowing for movement of the two contact points relative to one another (Fig. 4A) (12-16). The “cis” loop geometry is inaccessible in our double-tethered assays because the DNA is held in an extended configuration (Fig. 2B), which likely decouples loop extrusion from translocation. However, a cis loop configuration can be mimicked experimentally by providing a second DNA molecule in trans (Fig. 4B). To test the possible relationship between the observed linear translocation of condensin along the double-tethered DNA and the loop extrusion model, we asked whether condensin could move a second DNA substrate provided in trans relative to the tethered DNA. Indeed, fluorescently labeled (not extended) λ-DNA molecules added in trans moved at an apparent velocity of 66 ± 15 bp s-1 (16 ± 4 nm s-1, N=50) (Fig. 4C and D) while traveling an average distance of 14 ± 0.8 kb (3.4 ±0.3 μm, N=50) (Fig. 4E) – numbers that match excellently with the condensin motor properties measured above. These experiments strongly indicate that translocating condensin complexes were able to interact simultaneously with the tethered DNA and a second DNA, that condensin could translocate while bound to both DNA substrates, and that one piece of DNA was moved with respect to the other piece of DNA. We conclude that condensin is capable of moving two DNA substrates relative to one another, fulfilling a key expectation of the loop extrusion model.
Heretofore, a common argument against SMC proteins acting as molecular motors was their low rates of ATP hydrolysis when compared to other known nucleic acid motor proteins, which implied that they would not move fast enough to function efficiently on biologically relevant time scales. However, this discrepancy can be readily reconciled if condensin were able to take large steps, which is conceptually possible given its large 50-nm size. Based on the avaible data, it is in fact quite reasonable to invoke a large step size: Comparison of the single molecule translocation rate (∼60 bp s-1, or ∼14.9 nm s-1) to the bulk rate of ATP hydrolysis (Kcat = 2.0 s-1 in the presence of linear DNA) suggest that condensin may take steps on the order of ∼30 bp per molecule of ATP hydrolyzed. Even larger steps are inferred if each step is coupled to the hydrolysis of more than one molecule of ATP. These estimates assume that all of the proteins are ATPase active (one would deduce a smaller step size if a fraction of the protein were inactive), and also assume perfect coupling between ATP hydrolysis and translocation (while a more inefficient coupling would necessitate even larger step sizes). The idea that condensin takes very large steps is also consistent with the step sizes reported from magnetic tweezer experiments of DNA compaction induced by X. leavis condensin (80 ± 40 nm, Ref. (27) or S. cerevisiae condensin (J. Eeftens et al., unpublished). Such large step sizes would seem to rule out models for condensin movement similar to common DNA motor proteins such as helicases, translocases or polymerases, which are typically found to move in 1-bp increments (28-31). Higher-resolution measurements may prove informative for further defining the fundamental step size for translocating condensin.
To explain our novel results, we search for possible models for condensin motor activity that (i) can explain the relationship between a slow ATP hydrolysis rate relative to the rate of translocation; (ii) can accommodate a very large step size; and (iii) are consistent with the physical dimensions of the SMC complex. Based on these criteria, we can think of two theoretical possibilities, both of which use the SMC coiled-coils as the means of motility: Condensin might translocate along DNA through reiterative extension and retraction of the long Smc2–Smc4 coiled-coil domains, allowing for movement through a “scrunching” mechanism involving rod- to butterfly-like structural transitions (Fig. 4F); or condensin may perhaps use a myosin- or kinesin-like “walking” mechanism (Fig. 4G). The maximum single step size for each model is defined by the physical dimensions of the SMC coiled-coils, corresponding to ≲50 nm and ≲100 nm for the scrunching and walking mechanisms, respectively (Fig. 4F and G). Both models are consistent with the range of condensin architectures observed by electron microscopy and atomic force microscopy (21, 32). Movements might be powered by similar ATPase-dependent transitions between different structural states as reported for prokaryotic SMC complexes (33-35), though it remains to be determined how conformational changes could be translated into the directed movement depicted in our models. Further refinement of the translocation mechanism will depend upon fully defining the structural transitions that take place during the ATP hydrolysis cycle and establishing a better understanding of whether and if so, how different domains in the condensin complex engage DNA.
The finding that condensin is a mechanochemical motor capable of translocating along DNA has important implications for understanding fundamental mechanisms of chromosome organization across all domains of life. We propose that the ATP hydrolysis-dependent motor activity of condensin may be intimately linked to its role in promoting chromosome condensation, suggesting that condensin, and perhaps other SMC proteins, may provide the driving forces necessary to support 3D chromosome organization and compaction through a loop extrusion mechanism. These findings raise the question of whether other types of SMC complexes also exhibit intrinsic motor activity, and what molecular or regulatory features distinguish SMC motor proteins from those SMC complexes that seemingly lack motor activity.
Author contributions
T.T. designed and conducted single-molecule experiments and data analysis. S.B. purified condensin complexes, and conducted bulk biochemical measurements and electron microscopy analysis. J.E. designed and implemented single-molecule experiments. All authors discussed the experimental findings and co-wrote the manuscript.
Acknowledgements
We thank Damien D’Amours (Univ. of Montreal) for plasmids and yeast strains for overexpression of the condensin holocomplex, and members of the Haering, Greene, and Dekker laboratories for comments on the manuscript and the EMBL Electron Microscopy Facility and Proteomics Core Facilities for support. This work was funded by a MIRA grant from the National Institutes of Health to E.C.G. (R35GM118026), by EMBL and the ERC Consolidator grant to C.H.H. (ERC-2015-CoG 681365), and the ERC Advanced Grant SynDiv (ERC-ADG-2014 669598) and the Netherlands Organization for Scientific Research (NWO/OCW) (as part of the Frontiers of Nanoscience program) to C.D. T.T. was supported by a Japan Society for the Promotion of Science fellowship, and by a Uehara Memorial Foundation fellowship. S.B. was supported by an EMBL Interdisciplinary Postdoctoral fellowship (EIPOD) under Marie Curie Actions (COFUND). J.E. was supported by an EMBO short term fellowship.





{kind=link}
{kind=link}
{kind=link}
{kind=link}