

Abstract
Many animals are inhabited by microbial symbionts that influence their hosts’ development, physiology, ecological interactions, and evolutionary diversification. However, firm evidence for the existence and functional importance of resident microbiomes in larval Lepidoptera (caterpillars) is lacking, despite the fact that these insects are enormously diverse, major agricultural pests, and dominant herbivores in many ecosystems. Using 16S rRNA gene sequencing and quantitative PCR, we characterized the gut microbiomes of wild leaf-feeding caterpillars in the United States and Costa Rica, representing 124 species from 16 families. Compared with other insects and vertebrates assayed using the same methods, the microbes we detected in caterpillar guts were unusually low-density and highly variable among individuals. Furthermore, the abundance and composition of leaf-associated microbes were reflected in the feces of caterpillars consuming the same plants. Thus, microbes ingested with food are present (though possibly dead or dormant) in the caterpillar gut, but host-specific, resident symbionts are largely absent. To test whether transient microbes might still contribute to feeding and development, we conducted an experiment on field-collected caterpillars of the model species Manduca sexta. Antibiotic suppression of gut bacterial activity did not significantly affect caterpillar weight gain, development, or survival. The high pH, simple gut structure, and fast transit times that typify caterpillar digestive physiology may prevent microbial colonization. Moreover, host-encoded digestive and detoxification mechanisms likely render microbes unnecessary for caterpillar herbivory. Caterpillars illustrate the potential ecological and evolutionary benefits of independence from symbionts, a lifestyle which may be widespread among animals.
Introduction
Many animals are colonized by microbial symbionts that have beneficial and fundamentally important impacts on host biology. Microbes can regulate animal development, immunity and metabolism, mediate ecological interactions, and facilitate the evolutionary origin and diversification of animal clades (1–7). These integral host-microbe relationships have led to the notion that all animals can be conceptualized as “holobionts” (8–10), superorganism-like entities composed of the host plus its microbiome—defined here as the entire assemblage of commensal, pathogenic, and mutualistic microorganisms (11). Furthermore, the recent proliferation of microbiome surveys supports a widely held assumption that microbial symbioses are universal across animals (12, 13).
The Lepidoptera (butterflies, moths, and their caterpillar larvae), despite being key components of most terrestrial foodwebs and extraordinarily diverse (14), are one group in which the role of microbes remains ambiguous. Here we focus on caterpillars, which are the main—and in some Lepidoptera, the exclusive—feeding stage, and which have long been intensively studied in many fields (15). The vast majority of caterpillars are herbivores, and some insect herbivores rely on microbes to supplement missing nutrients, neutralize toxins, or digest plant cell walls (16, 17). However, considering caterpillars’ simple gut morphology and rapid digestive throughput, it has been speculated that microbes may be unable to persist in the caterpillar gut and do not contribute to digestion (18, 19). Indeed, microscopy-based studies report no, or minimal, microbial growth within the caterpillar gut (20–22).
DNA- and culture-based investigations of caterpillar gut microbiomes have produced mixed findings, with conflicting implications for microbial involvement in caterpillar biology. Some studies report a highly abundant and consistent bacterial community (23–25), characteristics that may indicate a functional association with the host. Others report high intraspecific variability in composition, and similarity between diet- and gut-associated microbes (26–29). Inconsistencies may arise from methodological factors such as contamination of low-biomass samples (30), starvation prior to sampling, sequencing of extracellular DNA (31), and the use of laboratory-raised insects or artificial diets (27, 32, 33). Regardless of these potential biases, when studying gut microbiomes, it is often difficult to distinguish between dead or dormant passengers (“transients” (34)) and persistent, living populations (“residents” (34) or “symbionts” sensu (35)). Further, microbes in the latter category may be parasitic or pathogenic, as well as beneficial. While microbes were known to cause disease in caterpillars as early as Louis Pasteur’s experiments on silkworms (36), their potential importance as mutualists remains unclear.
Do caterpillars depend on gut microbes for feeding and development? To answer this question, we surveyed microbiomes of a taxonomically and geographically broad array of wild caterpillars and conducted a field-based experiment on the model species Manduca sexta (Sphingidae). Our analyses are focused on the digestive tract, the most likely habitat for microbial colonization, as abundant microbes have not been observed elsewhere in the caterpillar body (32, 37). First, we characterized gut microbial abundance and composition across 124 species of actively feeding caterpillars in Costa Rica and the United States. We applied the same methods to 24 additional insect, bird, and mammal species that we expected to have functional microbiomes, to assess the reliability of our protocol and to contextualize our findings. Second, we experimentally tested whether gut bacteria impact the growth and survival of M. sexta. Our findings question the generality of animal-microbe symbioses, and may inform a multitude of research programs based on caterpillar herbivory in both natural and managed ecosystems (e.g., (38–41)).
Results
Survey of caterpillar gut microbiomes
Using quantitative PCR and sequencing of the 16S rRNA gene, we found that wild caterpillars representing a broad diversity of Lepidoptera had gut bacterial densities multiple orders of magnitude lower than the whole-body microbiomes of other insects and vertebrate feces measured using identical methods (p < 0.0001, Fig. 1A, Table S1). Some animals host symbiotic fungi (42), but fungal biomass was also lower in caterpillar guts relative to other insects and vertebrates (median 6.1 × 102 vs. 9.5 × 104 rRNA gene copies per gram, p < 0.0001). As another line of evidence of low microbial biomass, sequence libraries from caterpillar fecal samples were dominated by plant DNA. Though there was extensive variability, typically more than 80% of 16S rRNA gene sequences in caterpillar feces were from plant chloroplasts or mitochondria, versus 0.1% for other herbivores or omnivores with plant-rich diets (p < 0.0001, Fig. 1B). In a subset of caterpillars from which we sampled whole, homogenized midgut and hindgut tissue, plant DNA represented an even higher proportion of sequences in guts than in feces (Fig. S1A). This pattern is more likely a function of plant DNA degradation during intestinal transit than of bacterial proliferation, as bacterial density remained similar or decreased slightly from midgut to feces, depending on the caterpillar species (Fig. S1B).

Comparisons of bacterial densities, proportions of plant DNA in sequence libraries, and intraspecific variability between caterpillars and other animals expected to host functional microbiomes. Medians are indicated by black dashed lines, and points are horizontally jittered. Data for each species are listed in Table S1. One caterpillar species yielding <100 total sequences was excluded. A) The density of bacterial 16S rRNA gene copies in caterpillar feces versus fecal (vertebrates) or whole-body homogenate (other insect) samples of other animals (N=121 caterpillar species, 24 other species). Two caterpillar species with lower amplification than DNA extraction blanks are not shown. For species with multiple replicates, the median is plotted. B) The proportion of sequence libraries assigned to plant chloroplast or mitochondrial 16S rRNA (N=123 caterpillars, 21 other herbivores). C) The proportion of sequences belonging to core phylotypes, defined as those present in all conspecific individuals. Included are species with at least three replicates with >100 bacterial sequences each (N=7 caterpillars, 19 other animals). For species with more than three, points show the median core size across all combinations of three individuals, and error bars show the interquartile range.
Caterpillar gut bacterial assemblages also exhibited a high degree of intraspecific variability, as shown by higher beta diversity within caterpillar species relative to other insects and vertebrates (p = 0.0002). Such variability could indicate that the microbes found in caterpillar guts are generally transient, as animals with functionally important, resident microbiomes tend to host a high abundance of microbial taxa shared among conspecific individuals (e.g., (43–45)). In agreement with this expectation, within most species of the other animals analyzed here, microbiomes were largely made up of a common set of bacterial phylotypes. For example, >99% of sequences in any one honeybee belonged to phylotypes found in all honeybees included in the analysis. In contrast, even when raised on the same species of food plant under identical conditions, caterpillars had a much lower proportion of their gut bacterial assemblage belonging to core phylotypes (median 19.5%, p < 0.0001; Fig. 1C). In Schausiella santarosensis, which among caterpillars had the highest median core size of ∼50%, four of its six core phylotypes belong to Methylobacterium, a typical inhabitant of leaf surfaces (46). This observation hints that many of the core phylotypes which were found in caterpillars may be transient, food-derived microbes.
In addition to low total abundance and high inter-individual variability, caterpillar gut bacterial assemblages are dominated by leaf-associated taxa, further suggesting that resident, host-specific symbionts are sparse or absent. The bacterial phylotypes present in the feces of at least half of the sampled caterpillar individuals are Staphylococcus, Escherichia, Methylobacterium, Klebsiella/Enterobacter, Enterococcus, and Sphingomonas (Table S2). In Colorado and Costa Rica, we sampled leaf-associated bacteria from the same plant individuals consumed by the sampled caterpillars to examine whether leaves are a potential source of these taxa. Of the aforementioned phylotypes, all but Staphylococcus—a potential caterpillar pathogen (47) or, like Corynebacterium, a transient from human skin (48)—are also among the ten most common phylotypes found in leaf samples (Table S2). Across caterpillar individuals, a median 89.6% (interquartile range: 80.2–99.0%) of fecal bacterial sequences belonged to phylotypes detected on leaves. However, bacterial assemblages were not identical between leaves and caterpillar feces (p = 0.001). Besides the potential growth of parasites and/or mutualists in the gut, this difference could arise from digestion filtering out subsets of the leaf bacterial community.
Transient input of leaf-associated microbes could explain the substantial variation we observed in caterpillar gut bacterial loads (Fig. 1A). Leaf bacterial densities were highly variable within (tomato) and between (milkweed, eggplant, tomato) plant species, and this variation was reflected in the feces of monarch (Danaus plexxipus) and M. sexta caterpillars feeding on them (R2 = 0.24, p = 0.03; Fig. 2A). Furthermore, bacterial densities dropped by a median of 214-fold from leaves to feces, suggesting that any potential bacterial growth within the gut is relatively minor (Fig. 2A). The extent of this reduction varied widely (from 5 to 8400-fold, Fig. 2A), possibly because of inter-individual or interspecific differences in physiological traits that may eliminate leaf microbes, such as gut pH. As with patterns in total abundance, variation in bacterial taxonomic composition among leaves and caterpillar feces was correlated (Mantel r = 0.28, p = 0.001; Fig. 2B). In other words, caterpillars consuming leaves with more distinct bacterial assemblages produce more distinct bacterial assemblages in their feces, as would be expected from a digestive system in which microbes are diet-derived and only transiently present. Moreover, this process could explain a potential relationship between host relatedness and microbiome structure, a pattern sometimes interpreted to indicate functional host-symbiont interactions (49). Specifically, although confamilial caterpillars in Costa Rica had marginally more similar gut bacterial assemblages than did caterpillars in different families (p = 0.053), they had also been feeding on plants with especially similar leaf microbiomes (p = 0.005).

The abundance and composition of bacteria present in caterpillar fecal samples, as compared with paired diet (leaf) samples. A) The density of bacterial 16S rRNA gene copies in ground leaves versus feces, for 16 individuals collected in Colorado. Parallel lines indicate the association observed between plant and fecal bacterial abundances across paired samples. B) The correlation between beta diversity (Bray-Curtis dissimilarity metric) across caterpillar fecal samples collected in Costa Rica, and their paired leaf surface samples (N=24 caterpillar species, 19 plant species; 26 individuals each). Here only samples with >2,000 sequences are shown, to facilitate visualization.
Test of microbiome function in Manduca sexta
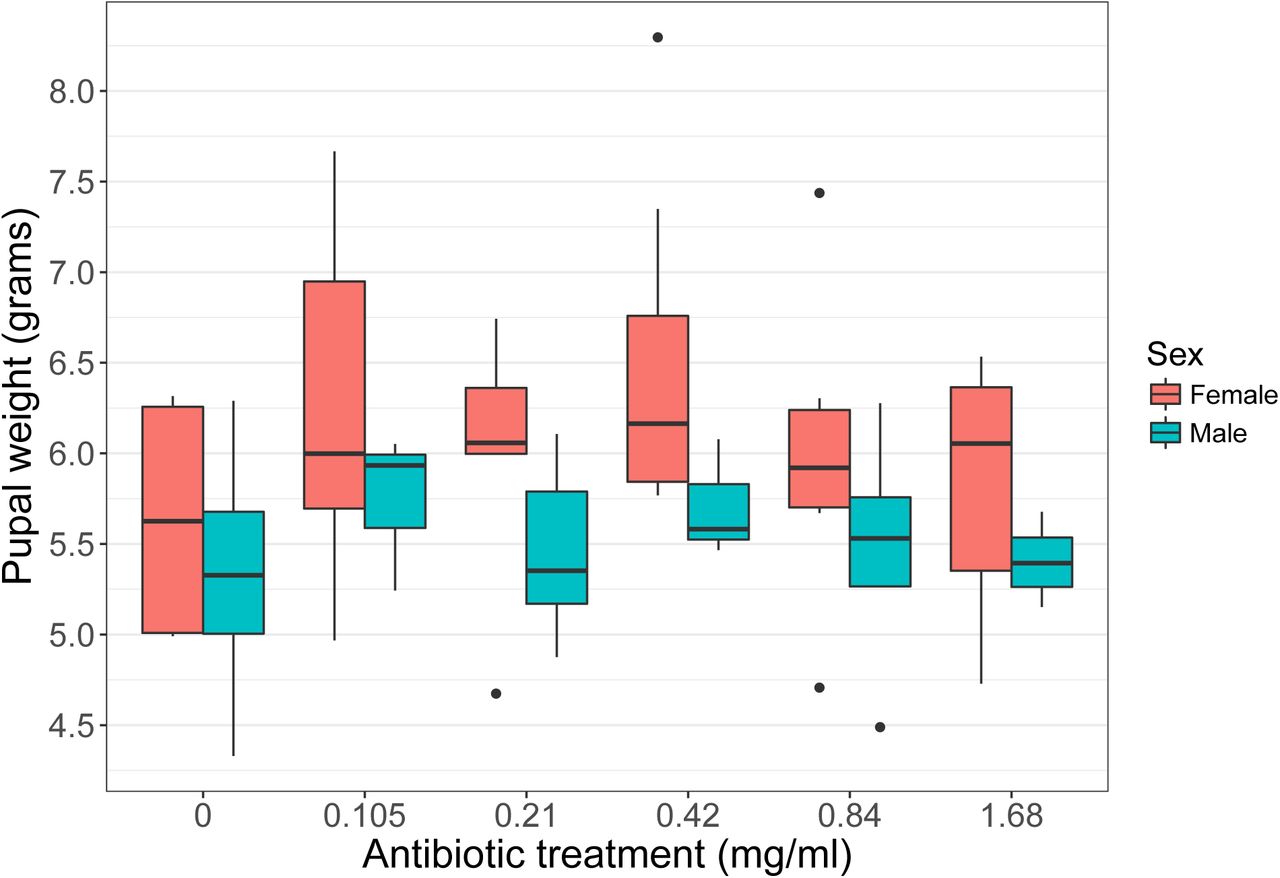
Supporting our claim that caterpillars lack resident, functional gut microbiomes, we show experimentally that the growth and survival of field-collected Manduca sexta caterpillars are not dependent on gut bacterial activity. As measured by qPCR, wild M. sexta contain ∼61,000-fold lower bacterial loads than expected from allometric scaling relationships based on animals with resident microbiomes ((50), Fig. S2). Feeding M. sexta antibiotics reduced this already low number of gut bacteria by 14- to 365-fold (range of medians across dosages), as measured using culture-dependent methods (R2 = 0.13, p = 0.003, Fig. S3A). These colony counts were positively correlated with the number of 16S rRNA gene copies (r = 0.38, p = 0.003; Fig. S3B). Suppression of viable bacteria had no effect on pupal weight (antibiotics: p = 0.45; sex: p = 0.014; interaction: p = 0.70; Fig. 3), which is correlated with fecundity in insects (51), nor on development time (antibiotics: p = 0.19; sex: p = 0.023; interaction: p = 0.63; Fig. S4A). Likewise, antibiotic treatment did not affect survival from larval hatching to adult emergence (p = 0.19, Fig. S4B), nor generally impact total feces production, which is an integrated measure of leaf consumption and assimilation efficiency (antibiotics: p = 0.07; sex: p = 0.002; interaction: p = 0.048). As expected with M. sexta (52) we found clear sexual size dimorphism, suggesting our experimental design had sufficient power to detect biologically meaningful differences. Given that antibiotics reduced fecal bacteria to a variable extent within and among treatments (Fig. S3A), we repeated the aforementioned analyses using gut bacterial abundance as the predictor variable. In all cases there was no significant relationship with host performance (p > 0.1), further indicating that reducing or eliminating gut bacteria from caterpillars does not negatively impact M. sexta fitness.

Increasing concentration of an antibiotic cocktail, delivered by a spray applied to Datura wrightii leaves prior to feeding, does not reduce Manduca sexta growth (N=62). Males and females are plotted separately, as they were expected to differ in size. Fresh weight was measured six days after pupation. Pupal weight correlates with adult fecundity and is often used as a proxy of insect fitness.
Discussion
Consistent with previous microscopy-based (20–22, 53) and molecular studies (26–29), we found that resident microbial symbionts are generally absent or present only in low numbers in caterpillar guts. As expected for herbivores consuming microbe-rich leaf tissue, diet-derived microbes are transiently present in caterpillar guts, wherein they may be dead or inactive. That the microbial biomass in caterpillar guts is far lower than in the guts or whole bodies of many other animals (Fig. 1A), and also lower than in their food (Fig. 2A), suggests a lack of persistent microbial growth within the gut. Moreover, any potential microbial metabolism might be too limited to substantially affect digestive processes, as illustrated by our observation that Manduca sexta caterpillars contain microbial loads orders of magnitude lower than comparably sized animals with resident microbiomes (Fig. S2). In addition to low abundance, the composition of microbes detected in caterpillar guts is highly variable among conspecific individuals (Fig. 1C). Lacking stable populations of core microbial taxa, caterpillar gut microbiomes may be easily influenced by the idiosyncrasies of which microbes are present on a given leaf and in what abundance, and which leaf microbes can survive transit through the digestive tract. Ingested microbes which die within the host may still be beneficial as a food source or by stimulating the immune system, but are not themselves symbionts (following the original definition of symbiosis as the “living together of different species” (referenced in (35)).
Based on the experiment with M. sexta, it is unlikely that microbes have cryptic, but essential, functions in caterpillar guts. Antibiotic suppression of viable gut bacterial loads in M. sexta had no apparent negative consequences, contrasting sharply with the many examples of major reductions in host growth or survival upon removal of beneficial symbionts (e.g., (54–56)). If anything, caterpillars treated with antibiotics showed slight (but not statistically significant) increases in performance (Fig. 3, Fig. S4B). Antibiotics increase the weight gain of laboratory-bred caterpillars (57–59), and commercially made caterpillar diets often contain antibiotics. This effect, also observed in livestock (60), might reflect microbial parasitism occurring in even apparently healthy caterpillars, and/or costly immune responses to the presence of pathogens (61). Aside from known leaf-specialists, some of the most frequently detected bacterial genera in this study (Table S2), including Acinetobacter, Clostridium, Enterobacter, Enterococcus, Escherichia, and Staphylococcus, have been reported to cause disease in caterpillars under some circumstances (37, 47, 62, 63).
The lack of a resident gut microbiome in caterpillars may directly result from a digestive physiology that is particularly unfavorable to microbial growth (18). The midgut, the largest section of the digestive tract wherein caterpillars digest leaf material and absorb the resulting nutrients (64), is a hostile environment for microbes (24). It is highly alkaline, with pH values often >10 (65) and as high as 12 (66), and contains host-encoded antimicrobial peptides (67). Additional attributes of the caterpillar gut that may hinder microbial colonization include a simple tube-like morphology without obvious microbe-housing structures (18), a continually replaced lining (the peritrophic matrix) covering the midgut epithelium (68) which may prevent biofilm formation, and short retention times (food transit takes ∼2 hours in M. sexta (69)). Although some insects harbor symbionts in specialized organs (53), to our knowledge, similar structures have not been reported in caterpillars. Buchner’s foundational survey of animal endosymbiosis describes Lepidoptera only as “a group in which no symbiont bearers have been discovered” ((53), p. 817). Moreover, previous studies did not find abundant microbes outside of the gut (32, 37).
Without the aid of microbial symbionts, how are caterpillars able to overcome the dietary challenges posed by herbivory? First, caterpillars use a combination of mechanical disruption, endogenously produced digestive enzymes, and high pH to extract easily solubilized nutrients, primarily from the contents of plant cells (18, 70, 71). Although this method of processing leaves is relatively inefficient, essential nutrients are not totally absent, so that caterpillars can compensate by simply eating more (18, 64). Some insects likely require microbes for detoxification (16), but many caterpillars possess host-encoded mechanisms for degrading or resisting plant allelochemicals (72). However, there may be a vestigial role for microbes in these processes, as genomes of many Lepidoptera contain microbial genes encoding enzymes with related functions (73, 74). These gene acquisitions may have enabled a symbiont-free feeding strategy.
The caterpillars surveyed here are likely to be representative of most externally leaf-feeding Lepidoptera, as we included a range of families, habitat types, and diet breadths from monophagous to highly generalist. However, a lack of resident gut microbiome in the caterpillar may not apply to the adult butterfly or moth. Compared with larvae, adult butterflies host distinct bacterial communities (32) and high gut microbial loads (75). Many other Lepidoptera lack functioning mouthparts or digestive tracts as adults, and in these groups microbes may be altogether irrelevant to digestion or nutrition. However, we cannot exclude the possibility that microbial symbionts may influence host fitness by their potential activities in eggs or pupae.
The extraordinary diversity and abundance of Lepidoptera (14) indicates that a symbiont-independent feeding strategy can be highly successful. Perhaps such success reflects a release from constraints imposed on other animals that do host and depend on symbionts. There are costs to engaging in mutualisms (e.g., (76–78)), and in a gut microbiome context one cost includes nutrient competition between host and microbes (60). A high availability of food allows caterpillars to “skim the cream” (64), assimilating simple nutrients that might otherwise be used by gut microbes and excreting recalcitrant material. In other words, “Why not do the digestion yourself rather than pay someone else to do it?” ((79), p. 53). Other costs include the risk of gut microbes becoming pathogenic (80, 81), and the potential for pathogens to exploit a gut environment that is hospitable to microbial mutualists. The extreme conditions in the caterpillar midgut may instead exclude all microbial growth, providing some degree of protection against disease.
Dependence on microbes with different physiological tolerances than the host constrains overall niche breadth (7, 77). As compared with groups lacking functional microbiomes, animals whose biology is heavily influenced by microbial mutualists may be less able to switch to new food plants or new habitats over evolutionary time. Indeed, it has been argued that while microbial symbioses can provide novel ecological functions, they may also increase the extinction risk of host lineages (7, 82). As Lepidoptera represent one of the most speciose animal radiations (83), a conspicuous question is whether independence from microbes may, in some cases, facilitate animal diversification.
Caterpillars do not appear to be unique in lacking a resident microbiome that is important for feeding and development. Microbiomes of walking sticks (84), sawfly larvae (85, 86), a saprophagous fly (87), a parasitic horsehair worm (88), a leaf beetle (89, 90), and certain ants (91) display features similar to those we observed in caterpillars. In fact, our data suggest that some vertebrates also have minimal gut microbiomes, and these species may feed relatively autonomously. Feces of the herbivorous brent goose (Branta bernicla) had low bacterial loads and a high proportion of plant DNA, and the insectivorous little brown bat (Myotis lucifugus) had similarly low fecal bacterial loads (Figs. 1A,B; Table S1). These species exhibit caterpillar-like physiological traits such as a relatively short gut and rapid digestive transit (92, 93). Additional examples in the microbiome literature might be obscured by contaminants masquerading as mutualists (94), a frequent lack of quantitative information (91) and experimental validation of microbial function in vivo, and publication bias against “negative results.”
While recent literature has documented extraordinary variation in the types of services provided by microbial symbionts, less explored is variation in the degree to which animals require any such services. Animals likely exist on a spectrum from tightly integrated host-microbe holobionts to simply animals, sensu stricto, in which a microbial presence is only relictual (i.e. mitochondria and horizontally transferred genes). Documenting the existence of microbially independent animals as well as their ecological, physiological and phylogenetic contexts, is a first step toward understanding the causes and consequences of evolutionary transitions along this continuum.
Methods
Sampling and Sequencing
Caterpillar fecal samples (N=185) were obtained from actively feeding, field-collected individuals in AZ, CO, MA, and NH, USA, and Área de Conservación Guanacaste, Costa Rica. To sample plant microbiomes, we collected leaves from the same branch used to feed caterpillars prior to fecal or gut sampling. Sequence composition was coreelated between feces and guts (Supplemental Methods), in line with a previous finding (32). All samples were preserved in 95% ethanol or dry at -20°C (90). We extracted DNA, PCR-amplified the 16S rRNA V4-V5 gene region and sequenced amplicons on an Illumina MiSeq in the same manner as previous insect microbiome studies (32, 90). These DNA extracts and primers were also used for quantitative PCR, which provides microbial biomass estimates concordant with those from microscopy (91) and culturing (Fig. S3B). We did not find evidence that low amplification of caterpillar fecal bacteria is due to primer bias, PCR inhibitors, or storage methods (Supplemental Methods).
Antibiotic Experiment
We collected Manduca sexta eggs from Datura wrightii plants near Portal, AZ, USA. 72 newly hatched larvae were randomly and evenly divided among six treatments varying from 0–1.68 mg total antibiotics per ml distilled water, and reared in separate unused plastic bags on D. wrightii foliage at the Southwestern Research Station. Water with or without antibiotics was sprayed onto leaves, which were briefly dried prior to feeding. The compounds used (rifampicin, tetracycline, streptomycin, in a 1:2:4 ratio) suppressed bacterial symbionts in other insect herbivores (55, 95). We collected a fresh fecal pellet from each caterpillar midway through the final instar, from which one subsample was cultured on LB media, and another used for qPCR and sequencing with the aforementioned protocol. Pupae were weighed six days after pupation and monitored daily for adult eclosion.
Data Analysis
Statistical analyses were conducted in R (96). Differences in bacterial loads, core sizes, and M. sexta performance variables were tested using linear models; residuals were visually inspected for Gaussian structure. The betadisper function in the vegan package was used to compare intraspecific beta diversity. M. sexta survival was analyzed using logistic regression. We used a Mantel test to estimate the rank correlation between leaf and fecal microbiome dissimilarities. A Wilcoxon test was used for proportions of plant DNA. Differences in community composition were analyzed using PERMANOVA. DNA sequences, metadata, and R code available at doi:10.6084/m9.figshare.4955648.
Acknowledgments
We thank the parataxonomists and staff of ACG and The Caterpillar Lab, without whom this project would not have been possible. J Dickerson, K Vaccarello, and H Layton provided invaluable help in the field and laboratory. HA Woods and MD Bowers aided in the planning and interpretation of this study. T Sharpton provided feedback on an earlier version of the manuscript. We also acknowledge administrative support from ACG, the Southwestern Research Station, and A Dietz. TJH was supported by the American Philosophical Society’s Lewis and Clark Fund, the National Science Foundation (NSF) Graduate Research Fellowship Program (1144083) and an NSF Doctoral Dissertation Improvement Grant (1601787). DHJ and WH were supported by the University of Pennsylvania, Wege Foundation, Permian Global, government of Costa Rica, and Guanacaste Dry Forest Conservation Fund.
References
- 1.↵
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.↵
- 8.↵
- 9.
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.
- 29.↵
- 30.↵
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.
- 40.
- 41.↵
- 42.↵
- 43.↵
- 44.
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.↵
- 86.↵
- 87.↵
- 88.↵
- 89.↵
- 90.↵
- 91.↵
- 92.↵
- 93.↵
- 94.↵
- 95.↵
- 96.↵





{kind=link}
{kind=link}
{kind=link}