

Abstract
Many aspects of neural physiology, including processes that underlie learning (e.g. neurotransmitter release and long-lasting changes in synaptic strength), are regulated by brief and local changes in [μm] levels of free intracellular Ca2+. On this scale, changes in [Ca2+] are known to activate many Ca2+-sensors, including the Ca2+/calmodulin-dependent kinases (CaMKs). Although CaMK4 is known to function in long-term memory and its paralog, CaMK1, in nervous system development, there is no evidence indicating that they function in learning acquisition. Here we reveal that the Caenorhabditis elegans ortholog of CaMK1/4, CMK-1, regulates responses to mechanical stimuli and learning, specifically habituation, a conserved form of non-associative learning. The habituation phenotypes of cmk-1 mutants are sensitive to interstimulus interval (ISI), such that cmk-1 mediates habituation rate at short ISIs and habituation level at long ISIs. This is the first in vivo evidence that CaMK1/4 functions to modulate learning acquisition in awake, behaving animals and provides direct evidence to support the hypothesis that different mechanisms mediate habituation learning at different ISIs. From catalytic site analysis of the human and C. elegans CaMKs, we predicted potential CaMK phosphorylation targets and, through mutation studies, identified one of these, O-linked N-acetylglucosamine (O-GlcNAc) transferase, OGT-1, as also being necessary for wild-type responses to mechanical stimuli and learning. Detailed behavioral analysis of single and double mutants suggests that CMK-1 and OGT-1 function in parallel pathways that may converge on a common substrate to modulate the tap response. Our results provide the first evidence of a role for CaMK and O-GlcNAc post-translational modification in responding to mechanical stimuli and learning, which are fundamental biological processes present in all animals.
Introduction
Learning is a fundamental biological process by which organisms update and modify their behavioral output to sensory stimuli based on past experience. This important phenomenon allows organisms to adapt their behavior to best suit the current conditions in the inconstant environment. Brief and local changes in micromolar concentration levels of free intracellular Ca2+ play a critical role in modifying many aspects of neural physiology such as learning. Changes in [Ca2+] on this scale are known to activate many Ca2+-sensors, including the important and ubiquitously expressed protein calmodulin (CaM). Once bound with four Ca2+ ions, CaM (Ca2+/CaM) is known to regulate many different signaling proteins, including the CaM-kinase (CaMK) family of protein-serine/threonine kinases, which are highly expressed in the nervous system. Within the larger CaMK group (consists of ~ 23 kinase families) the CaMK1 family (consisting of CaMKK, CaMK1 and CaMK4; reviewed in 1) has been shown to play important roles in the context of nervous system development and plasticity.
CaMKK is purported to function upstream of both CaMK1 and CaMK4 by phosphorylating Thr residues in the activation site of these kinases to increase their Ca2+/CaM-dependent phosphotransferase activities (2, 3). The substrate recognition motifs of numerous protein kinases have significant overlap and correct target specificity is thought to also be regulated by localization of the kinase and its substrates within the cell (4); this has been established for CaMK1 and CaMK4 (5).
CaMK4’s expression is usually restricted to the nucleus (6-8), where it has been shown to regulate gene transcription during the induction of long-term synaptic plasticity and long-term memory in rodents (9-11). In contrast, CaMK1’s expression has been observed to be cytoplasmic in most cases (12). Many studies have demonstrated an important role for CaMK1 in the developing mammalian nervous system, specifically regulating axonal growth cone motility and axonal outgrowth (13), dendritic arborization (14-16), and formation of dendritic spines and synapses (17).
More recent studies have attempted to ascertain whether CaMK1 plays a role in plasticity of the nervous system; Schmitt et al. (18) investigated the role of CaMKK, CaMK1 and CaMK4 in long-term potentiation (LTP) and demonstrated that CaMKK and CaMK1, but not CaMK4, play a role in activating Ras-extracellular signal-regulated protein kinase (Ras-ERK) signaling during early-phase LTP in hippocampal neuron cultures. Using the same experimental preparation, Guire et al. (19) went on to demonstrate that CaMK1 also functions during LTP to recruit calcium permeable AMPA receptors. These studies indicate that similar to CaMK4, CaMK1 might also function in plasticity and perhaps learning and memory, but unlike CaMK4 it most likely functions near the synapse and in shorter forms of plasticity. To date, no one has directly tested whether CaMK1 plays a role in plasticity or learning and memory in any organism in vivo.
The nematode Caenorhabditis elegans responds to a non-localized mechanosensory stimulus, a tap to the side of the Petri plate it inhabits, by performing a reversal (changing from forward to backward locomotion). In wild-type worms, repeated administration of the tap stimulus results in habituation, a form of non-associative learning, which can be observed as a decrease in both the size of the reversal (response magnitude) and the likelihood of responding (response probability) (20, 21).
Ca2+ has been shown to be an important modulator of tap habituation in C. elegans (22, 23). Repeated mechanical stimulation resulted in an attenuation of the Ca2+ transient that followed each mechanical stimulus (23). Chelation of the tap-induced Ca2+ transient resulted in more rapid habituation, as did mutations in the genes encoding the calcium signaling molecules calreticulin (crt-1) and the inositol triphosphate receptor (itr-1: 22). Despite the obvious importance of Ca2+ signaling in this phenomenon, components of the CaMK cascade have yet to be tested for their role in habituation. The catalytic activity of proteins in this signaling cascade are dependent upon increases in intracellular Ca2+ concentration (which occurs in response to tap), are highly expressed in the nervous system, and have been shown to be important for cellular models of plasticity and long-term memory (reviewed in 24). Thus we hypothesized that this cascade may also function in learning in C. elegans, specifically in tap habituation.
The C. elegans genome encodes a single homolog each of CaMKK, ckk-1, and CaMK1/4, cmk-1 (25). Both genes are expressed in the nervous system and strains with mutations in these genes appear superficially wild-type (26), however several exciting studies have recently characterized molecular mechanisms underlying behavioral deficits related to experience-dependent thermotaxis and heat avoidance (27; 28; 29). We found that C. elegans strains carrying mutations in cmk-1, but not ckk-1, exhibited larger responses to mechanical stimuli and a higher final habituated level of responding than wild-type animals when stimuli were presented every sixty seconds. Intriguingly, the initial response and final habituated level phenotypes of cmk-1 mutants were genetically dissociable and sensitive to the interstimulus interval (ISI) at which the mechanosensory stimuli were presented. This is the first in vivo evidence that CaMK1/4 functions to modulate learning acquisition in awake, behaving animals and provides direct evidence to support the hypothesis that different mechanisms mediate habituation learning at different ISIs (Rankin and Broster, 1992). A screen for downstream targets of CMK-1 predicted from bioinformatics analysis of the human and C. elegans CaMK catalytic domains led to the identification of the C. elegans O-linked N-acetylglucosamine (O-GlcNAc) transferase homolog, OGT-1, as also functioning in responding to mechanical stimuli and tap habituation. Thus, we demonstrate novel roles for posttranslational O-GlcNAc modification of proteins in responding to mechanical stimuli and in vivo learning.
Results
CMK-1 Modulates the Rate of Habituation at Short Interstimulus Intervals (10s) and the Final Habituated Level of Responding at Long Interstimulus Intervals (60s)
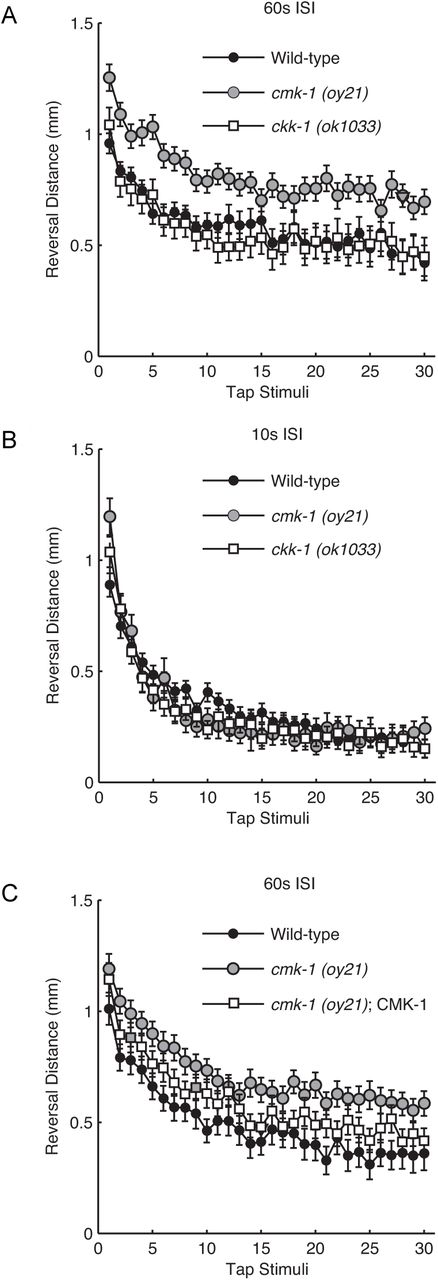
To determine whether CaMKs function in learning we examined habituation to repeated tap stimuli by the C. elegans putative null mutants of mammalian CaMK1/CaMK4, cmk-1(oy21), and CaMKK, ckk-1(ok1033), homologs. Wild-type worms habituate to repeated tap stimuli by decreasing the distance they reverse (20, 21). ckk-1(ok1033) mutant worms responded initially and habituated similarly to wild-type worms at both 10 and 60 s ISIs (pNS for all, Fig. 1A-B). In contrast, cmk-1 mutant worms displayed larger initial responses to tap (p<0.001, Fig. 1A) and habituation deficits dependent on the rate at which stimuli were delivered. If stimuli were administered at a 60 s ISI, a larger tap response was apparent across training (p<0.001; Fig. 1A), however if stimuli were presented every 10 s, the cmk-1 mutant habituated faster (greater rate of response decrement) than wild-type, but to an asymptotic level indistinguishable from wild-type (pNS; Fig. 1B). Therefore, the elevated final reversal distance habituation level in cmk-1 mutants (Fig. 1A) is not caused by a general increase in responding, but instead is specific to the stimuli being delivered at a 60s ISI. Wild-type habituation to tap stimuli presented at a 60s ISI was restored in cmk-1(oy21) mutant worms expressing wild-type CMK-1 cDNA under control of its endogenous promoter (N2 vs. rescue pNS; Fig. 1C); confirming oy21 as the causative allele.

Response to mechanical stimuli and tap habituation of wild-type, cmk-1(oy21), and ckk-1(ok1033) mutants. (A) Reversal distance habituation to tap stimuli presented at a 60s ISI. (B) Reversal distance habituation to tap stimuli presented at a 10s ISI. (C) Response to mechanical stimuli and tap habituation of wild-type, cmk-1(oy21), and genetic rescue with CMK-1 cDNA in cmk-1(oy21) mutants. Error bars represent 95% confidence intervals (Clopper-Pearson method for binomial data).
Age is an important factor that modulates habituation (30), but the habituation phenotype of cmk-1 mutants was unlikely to be due to differences in growth or aging, as the mutant worms began egg-laying at a similar time as wild-type worms (data not shown). Additionally, the difference in habituation between cmk-1 mutant and wild-type worms at a 60s ISI was present regardless of which day of adulthood the worms were tested on (p<0.05; Fig. S1 and S2). Note that in younger adults, the initial responses are of normal size, but the habituated responses are larger (Fig. S1), dissociating the initial and final response metrics. This further suggests that cmk-1 modulates habituation in an ISI-dependent manner, as the cmk-1 mutant displayed an altered final habituated level of responding at a 60s ISI and altered rate of habituation at a 10s ISI, even at an age where the increased initial response phenotype of cmk-1 mutants is not present (Fig. S1).
Loss of CMK-1 phosphorylation site (T179) recapitulates null allele
Although previous studies suggested that CaMKK stimulates the kinase activity of CaMK1/4, our results indicate that CMK-1, but not CKK-1, functions in habituation. Consistent with this assertion, the expression pattern of cmk-1 is broader than ckk-1 (26) and we found that cmk-1, but not ckk-1, was expressed in the touch receptor neurons and interneurons of the tap withdrawal circuit (Fig. S3). This suggests that either (i) activation of CaMK by calmodulin alone may be sufficient to activate the kinase in the context of some biological signaling, and/or (ii) CaMK is activated via phosphorylation by another unidentified kinase. To test these hypotheses we took advantage of a cmk-1 point mutant (gk691866) whose conserved CaMKK phosphorylation site, Threonine-179 (T179; 2, 3), was mutated to an isoleucine (I) and could therefore no longer be phosphorylated. We performed a complementation test between the cmk-1(gk691866) point mutant, T179I, and the cmk-1(oy21) null mutant and reasoned that if these alleles complemented in the context of habituation it would indicate that phosphorylation of T179 was not required for wild-type habituation. Interestingly, we observed that compared to wild-type worms and cmk-1(oy21/+) heterozygotes, cmk-1(oy21/gk691866) heterozygote mutants showed significantly increased responding to the initial tap (p=0.05 and p<0.001, respectively) and a higher final level of habitation to tap stimuli (p<0.01 and p<0.001, respectively); indicating that these alleles failed to complement (Fig. 2). Thus, these data support a role for phosphorylation of T179 as being necessary for wild-type responding to mechanical stimuli and habituation. Furthermore, because we demonstrated above that CKK-1 does not function in habituation, CMK-1 must be activated via T179 phosphorylation by another, as yet unidentified, kinase.

Response to mechanical stimuli and tap habituation of wild-type, cmk-1(oy21), cmk-1(oy21/+), and cmk-1(oy21/gk691866) worms when stimuli are presented at a 60s ISI and reversal distance is measured. Error bars represent 95% confidence intervals.
Genetic dissociation of initial response and habituation phenotypes
In mammals, the orthologs of CMK-1, CaMK1 and CaMK4, localize to different subcellular compartments (the cytoplasm and the nucleus, respectively). Consistent with previous reports (Schild et al., 2014), we found that CMK-1::GFP fusion proteins localize to the cytoplasm in cell bodies and neurites and are largely excluded from the nucleus (Fig. 3A). In a screen for noxious heat avoidance defects, Schild et al. (2014) isolated a cmk-1 gain-of-function allele, pg58, which encodes a truncated protein lacking most of its regulatory domain and a nuclear export sequence (NES), but with an intact kinase catalytic domain. As had been observed with a similar mutation in mammalian CaMKI (Stedman et al., 2004), Schild et al. (2014) found that truncated CMK-1(1-304) abnormally accumulated in the nucleus. In their model, CMK-1 shuttles between cytoplasm and nucleus to modulate noxious heat avoidance. To test whether a similar process mediated habituation to tap, we evaluated learning in the cmk-1(pg58) mutant. Although they had a large initial response to tap (p<0.05; Fig. 3B) the final habituated response size of cmk-1(pg58) was indistinguishable from wild-type (pNS; Fig. 3B), suggesting appropriate subcellular localization is essential for setting naïve responsivity to tap, but not necessarily modulating it. By dissociating the initial response and habituation phenotypes of the cmk-1 mutant, results from this allele confirm they are independent metrics.

(A) CMK-1::GFP fusion proteins localize to the cytoplasm in cell bodies and neurites and are largely excluded from the nucleus. (B) Reversal response distance and habituation of wild-type, and cmk-1(pg58) mutants to repeated tap stimuli presented at a 60s ISI.
Identification of Candidate CMK-1 Target Proteins Through Analysis of Evolutionarily Conserved Predicted CaMK Phosphosites
Protein kinases recognize the specific Ser/Thr/Tyr amino acid residues that they phosphorylate on their target substrates based upon the sequence of residues that flank the phosphoacceptor site (comprises the kinase consensus sequence). This is due to kinase-substrate binding following a lock and key model, whereby the peptide sequence flanking the phosphosite on the target protein fits into the catalytic domain of the kinase, because of the presence of specificity-determining residues located there which often directly interact with the side chains of amino acid sequences surrounding phosphosites in substrates (32). These principles have been previously used to predict the kinase substrate specificities of 492 human protein kinases in silico (33).
The computational methods have now been further improved with refinements of the original algorithms and training data from over 10,000 kinase-protein phosphosite pairs and 8,000 kinase-peptide phosphosite pairs. We used these updated methods to generate a kinase substrate specificity prediction matrix (KSSPM) for C. elegans CMK-1 based on the primary amino acid sequence of its catalytic domain, and then used this KSSPM to query all of the 20,470 known C. elegans protein sequences to identify those proteins that featured the top 600 highest scoring predicted phosphosites (Table S1). Next, we identified the closest human cognate proteins that featured similar phosphosites, and then scored the human phosphosites with KSSPMs for all four human CaMK1 isoforms and CaMK4 (which share 65% and 44% sequence identity, as measured by Blastp, with the C. elegans CMK-1 protein, respectively; Fig. S4). Of particular interest were those C. elegans protein and phosphosites that were highly conserved in Homo sapiens and predicted to be targeted by human CaMK1 isoforms and CaMK4. Such high evolutionary conservation would support important functional roles for these kinase-substrate pairs. More information about the predicted phosphorylation of these human phosphosites by human protein kinases and their evolutionary conservation in over 20 other species is available in the PhosphoNET website at www.phosphonet.ca.
The above-generated list of 600 phosphosites in 373 C. elegans proteins predicted to be CMK-1 targets was used to prioritize candidates. Candidates that had been previously shown to interact with CaMKs for which testable knockout mutant alleles were available were of highest interest, as well those ranked within the top 20 by p-site score and assayed them for habituation at a 60 s ISI. Out of the 22 mutants tested to date, 17 were observed to show an initial response and/or habituation phenotype (Fig. S5, S6 and (Table 1). A Venn diagram was generated that grouped genes that when mutated showed similar behavioral phenotypes (Fig. 4).

Venn diagram grouping genes whose mutant alleles showed similar behavioral phenotypes when given 30 taps at a 60s ISI. Grouping was based on the statistical analysis presented in Table 1.
Summary of mechanosensory and learning phenotypes for predicted CMK-1 phosphorylation targets.
O-GlcNAc transferase, OGT-1, Functions in Habituation and is Expressed in the Nervous System, including the Touch Receptor Neurons
ogt-1 mutants showed an initial mechanosensory response and habituation phenotype that was strikingly similar to cmk-1 mutants; ogt-1(ok430) mutants were significantly more responsive to the initial tap (p<0.001), and showed significantly higher final level of habituation to stimuli delivered at a 60s ISI (p<0.001; Fig. 5A, Table 1). Because of this we further investigated the role of this protein in habituation by assaying whether OGT-1 was also similar to CMK-1 in its ISI dependency (i.e. whether it mediated distinct aspects of habituation at a 10s ISI versus a 60s ISI). When we habituated ogt-1(ok430) mutants at a 10s ISI instead of a 60s ISI, we observed that they were still more responsive to the initial tap (p<0.001), but they habituated more rapidly and to the same final level as wild-type worms in the measure of reversal distance (pNS; Fig. 5B). Thus, ogt-1 mutants robustly phenocopy cmk-1 mutants in all measures tested. Importantly, Hanover et al. (34) has demonstrated that the ogt-1(ok430) allele is a true null and results in the complete loss of function of this transferase; in ogt-1(ok430) mutants O-glycNAc is absent. This was shown via direct measurement of O-GlcNAcitol released after alkaline β-elimination and by O-GlcNAc antibody staining (34).

Response to mechanical stimuli and tap habituation of wild-type, and ogt-1(ok430) mutants. (A) Reversal distance habituation to tap stimuli presented at a 60s ISI. (B) Reversal distance habituation to tap stimuli presented at a 10s ISI. (C) Mechanosensory neurons, ALMs and AVM, (visualized with Pmec-7::mRFP) express OGT-1 (visualized with Pogt-1::GFP). Anterior is at left
We next confirmed that the mutation in ogt-1, ok430, was indeed the mutation which caused the mechanoresponding and habituation phenotypes observed above by testing a second null allele of ogt-1, tm1046. Tm1046 is a 466 bp deletion, resulting in a frameshift and an early stop after 392 amino acids. ogt-1(tm1046) mutants displayed an initial response (p<0.001) and habituation phenotype (p<0.001) similar to ogt-1(ok430) null mutants (Fig. 5; data not shown), thus demonstrating that OGT-1 function is critical for wild-type responses to tap and habituation.
To evaluate the expression pattern of ogt-1 we created a transcriptional reporter that consisted of ~2 kb of the ogt-1 promoter fused to GFP (Pogt-1 ::GFP) and injected this into wild-type worms. Imaging of this reporter revealed that OGT-1 is expressed quite broadly across the nervous system, including the touch cells, in addition to muscles and seam cells (Fig. 5C).
cmk-1 and ogt-1 Exhibit a Complex Genetic Interaction
To test whether cmk-1 and ogt-1 interact genetically we assayed the habituation of cmk-1(oy21); ogt-1(ok430) double mutants. When these double mutants were habituated at a 60s ISI they acted additively to give significantly larger initial responses (p<0.001 for both) and larger reversal distance habituation phenotypes than that of either single mutant (p<0.01 for both; Fig. 6A). The most parsimonious explanation of this data is that each mutation contributes to the habituation phenotype through disruption of independent genetic pathways. This result was surprising given that the individual mutant phenotypes were so strikingly similar and that the mammalian homologues of these proteins have been shown to interact (35, 36). Because the response distance variable is actually composed of two different measures, reversal speed and reversal duration, we decided to further analyze their relationship by breaking reversal distance down into these two components and examining their habituation (Fig. 6B and 6C).

Response to mechanical stimuli and tap habituation of wild-type, cmk-1, ogt-1 and cmk-1;ogt-1 double mutants when stimuli are presented at a 60s ISI. (A) Reversal distance habituation to tap stimuli. (B) Reversal speed habituation to tap stimuli. (C) Reversal duration habituation. Error bars represent 95% confidence intervals.
For the initial response to tap, the cmk-1 single mutant exhibited significantly faster reversals (Fig. 6B) of increased duration (Fig. 6C) compared to wild-type. In contrast, the ogt-1 mutant reversed at a speed similar to wild-type (Fig. 6), but with a duration longer than even the cmk-1 mutant (Fig. 6C). Thus, the single mutant data indicate that the similar reversal distance phenotype of unhabituated cmk-1 and ogt-1 mutants was caused by distinct processes, with CMK-1 primarily influencing speed of reversals and modestly influencing reversal duration, and OGT-1 influencing duration, but not speed. However, cmk-1; ogt-1 double mutants performed significantly slower reversals than cmk-1 single mutants (p<0.001), almost indistinguishable from wild-type (p=0.04), indicating that ogt-1 may act as a suppressor of cmk-1 for reversal speed in naïve animals (Fig. 6B). Thus, in the context of responding to an initial mechanical stimulus, there appears to be some interaction between the cmk-1 and ogt-1 pathways.
In the case of habituated animals, both cmk-1 and ogt-1 single mutants exhibited significantly faster reversals compared to wild-type and a consistent, but not significant trend towards increased duration, phenotypes which were additive, giving the rapid and long-lasting final responses of the cmk-1; ogt-1 double mutant.
Discussion
We report here that the C. elegans CMK-1, but not CKK-1, mediates habituation to mechanosensory stimuli in an ISI-dependent manner. Through catalytic site analysis of CaMKs we predicted and screened potential CaMK phosphorylation targets to identify the null OGT mutant, ogt-1, as also functioning in this form of learning. We showed that similar to cmk-1, ogt-1 mutants altered habituation in an ISI-dependent manner. Finally, we assayed the tap habituation of cmk-1(oy21); ogt-1(430) double mutants and found that these two genes have a complex genetic interaction in the context of naïve responding, yet exhibit an additive habituation phenotype.
While many previous studies have suggested that CaMK1/4 and CaMKK can function in the same pathway, our results indicate that CMK-1, but not CKK-1, functions in responses and habituation to mechanical stimuli are not the first findings to indicate that CaMK1/4 can function independently of CaMKK. In C. elegans, Kimura et al. (26) found that CMK-1 is expressed in more neurons than is CKK-1 and that although CKK-1 enhanced the CMK-1- dependent phosphorylation of a transcription factor (CREB), CMK-1 was able to phosphorylate CREB in the absence of CKK-1. Similarly, Satterlee et al. (37) found that CMK-1, but not CKK-1, functioned to regulate AFD sensory neuron specific gene expression in C. elegans. These findings also appear to be consistent with studies in vertebrate organisms where CaMK1 is known to have a wider expression profile than CaMKK (reviewed in 38). These data indicate that either in some cases (i) activation of CaMK1/4 by calmodulin alone may be sufficient to activate the kinase in the context of some biological signaling or (ii) CaMK1/4 is activated via phosphorylation by another unidentified kinase. Although these possibilities are not necessarily mutually exclusive, our results that showed that the cmk-1(gk691866) point mutant, T179I, and the cmk-1(oy21) null mutant fail to complement in the context of naïve mechanosenory responding and habituation indicate that, at least in this organism and these phenotypes, CMK-1 is activated via phosphorylation by another as yet unidentified kinase. Future work is aimed at identifying the cellular and subcellular site of action for CMK-1 and OGT-1. A mutant with mislocalized CMK-1 responded initially with large reversals, but habituated normally, suggesting a shuttling of CMK-1 into the nucleus does not mediate tap habituation, as it does heat avoidance (28).
To find downstream targets of CMK-1 in C. elegans we used a novel bioinformatics approach to generate a list of protein candidates predicted to be phosphorylated by CMK-1 to screen for tap response and habituation phenotypes, similar to those observed in cmk-1 mutants. Our bioinformatics approach to generate a list enriched in proteins that were likely to function in either responding to mechanical stimuli and/or mechanosensory habituation used an effective kinase substrate prediction algorithm developed by Safaei and colleagues (33). Behaviorally screening strains with mutations in our top candidates, we found that this list was indeed enriched for mutations which cause tap response and/or habituation deficits, as 17/22 strains deviated from wild-type in some aspect of these behaviors. Thus this approach appears to be an effective method to identify downstream candidates for phosphorylation targets of kinases in the context of mechanosensory responding and learning.
By screening knockout mutants from among the top candidates of the CaMK phosphosite substrate prediction, we found that the C. elegans O-GlcNAc transferase (OGT) mutants, ogt-1, displayed strikingly similar phenotypes compared to cmk-1 mutants. O-GlcNAc glycosylation is a unique and dynamic cytosolic and nuclear carbohydrate post-translational modification in which β-N-acetylglucosamine is covalently attached to serine or threonine residues of proteins. In contrast to other forms of glycosylation, O-GlcNAc glycosylation occurs intracellularly, is rapid (occur as quickly as 1-5 min after cellular stimulation; 35, 39) and is not further modified into complex glycans. Hence, it is thought to be more akin to phosphorylation than to other forms of glycosylation. Interestingly, O-GlcNAc glycosylation sometimes occurs on or near serine or threonine residues that are also known to undergo phosphorylation (reviewed in 40). Both OGT and O-GlcNAcase (enzyme that functions in opposition to OGT to remove β-N-acetylglucosamine residues; OGA) are highly expressed in the brain (41, 42) and enriched at synapses within neurons (43,44).
In C. elegans, although OGT-1 is expressed in the embryonic nervous system (45) no role has yet been described for this protein in responding to mechanosensory stimuli or learning. Instead, studies have shown that it functions in macronutrient storage (34), dauer formation (46), lifespan (47), the glucose stress response (48) and proteotoxicity in C. elegans neurodegenerative disease models (49). In other species a role for O-GlcNAc glycosylation has yet to be demonstrated in vivo for learning acquisition, but it has been shown to function in long-term memory (50) and in cellular models of plasticity such as: long-term potentiation (LTP), long-term depression (51) and paired-pulse facilitation (52). Thus, our evidence of an in vivo requirement for OGT-1 in learning in C. elegans will very likely be conserved in learning in other species, including mammals.
Epistasis experiments measuring reversal distance revealed the cmk-1(oy21); ogt-1(430) double mutant phenotype to be additive for both naïve and habituated states. Additivety is most often thought to be suggestive of a sign of the absence of a functional relationship between the mutated genes under study but in some cases it may be due to both gene products converging on the same downstream target(s), and/or non-linear dynamics of regulatory networks (53).
Although, clearly, the most parsimonious explanation for the additive genetic interaction between null mutations in cmk-1 and ogt-1 is that each mutation contributes to the habituation phenotype through disruption of at least two contributing independent linear biochemical pathways; data from previous studies have revealed that there is much complex cross talk between O-GlcNAc and phosphorylation signaling, and thus it is tempting to speculate that the relationship between CMK-1 and OGT-1 may be more intimate. Given that (i) cmk-1 and ogt-1 mechanosensory responding and two different habituation phenotypes were so strikingly similar, (ii) mammalian CaMK4 and OGT interact (CaMK4 phosphorylates OGT (35), and OGT can add O-GlcNAcs to CaMK4 (36)), (iii) mammalian CaMK4 and OGT are known to share several common substrates (CREB, serum response factor, the CREB-binding protein, and OGT itself; 36, 54, 55-57), and (iv) virtually every other O-GlcNAcylated protein identified (> 250) has also been shown to be a phosphoprotein (i.e. it is phosphorylated by a kinase; 58, 59) we hypothesized that the additive phenotypes of cmk-1; ogt-1 double mutants may be caused by CMK-1 and OGT-1 sharing a downstream substrate and/or functioning in a non-linear genetic pathway.
To test this hypothesis, we further analyzed this genetic interaction by separating reversal distance into its two components, reversal speed and reversal duration, for both naïve and habituated mechanosensory responses. The clearest conclusion can be drawn from the double mutant data in the context of responding to tap stimuli. When naïve reversal speed is measured, cmk-1; ogt-1 double mutants perform significantly slower reversals than cmk-1 single mutants, very similar to wild-type, indicating that ogt-1 acts as a suppressor of cmk-1. Because the cmk-1(oy21) allele used in these experiments is very likely a null allele, these data support the existence of ogt-1 functions downstream of cmk-1 to negatively regulate the pathway in wild-type animals. When naïve reversal duration is measured, the cmk-1; ogt-1 double mutant phenotype is synergistic. Synergy in double mutants affected in two non-homologous genes can be a consequence of genes disrupting steps of pathways that converge at some node of a regulatory network, such as sharing a common substrate (reviewed in 60), suggesting that CMK-1 and OGT-1 share a common downstream target(s).
Additionally, a synergistic phenotype is also observed for the baseline locomotory speed in cmk-1; ogt-1 double mutants measured before mechanosensory stimuli were administered (Fig. S7); providing additional phenotypic support to our hypothesis that these genetic pathways intersect. In the context of habituation, the additive habituation phenotypes regardless of the measure in cmk-1; ogt-1 double mutants indicate that in this context cmk-1 and ogt-1 function in two parallel pathways, which may converge on a common downstream substrate.
Using all available published data on CaMK1/4 and OGT as well as from the experiments performed in this study, we have a devised a model for this non-linear genetic network in C. elegans (Fig. 7). Biochemical studies, outside of the scope of this work, will be required to validate this hypothesis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proposed functional model that explains the genetic interaction between cmk-1 and ogt-1 mutants. Model aspects supported by data from this study are represented in blue.
To our knowledge, cmk-1 and ogt-1 represent the first reported mechanosensory hypersensitive mutants in C. elegans despite several pioneering genetic screens characterizing harsh and soft touch mechanosensory defective mutants by Chalfie and colleagues (61,62). Further, we have shown that cmk-1(pg58) mutants also display mechanosensory hyperesponsivity, suggesting that wild-type shuttling of cmk-1 between the cytoplasm and the nucleus is necessary for setting response level to mechanical stimuli – similar to it’s effects on thermotaxis and heat avoidance (27, 28, 29, 37). Further characterization of cmk-1 interacting proteins may identify additional mechanosensory hypersensitive mutants, an as yet largely uncharacterized family of genes regulating mechanosensation.
Finally, our findings identify the first proteins that specifically alter habituation differently at different ISIs: CMK-1 and OGT-1. Early detailed parametric studies of C. elegans mechanosensory habituation found that the rate and asymptotic level of habituation differed depending on what ISI stimuli were presented; mechanosensory stimuli which were presented at a 10s ISI resulted in faster and deeper habituation than stimuli presented at a 60s ISI (20). These experiments led to the hypothesis that habituation at different ISIs may recruit/require different molecular mechanisms. Our findings that cmk-1 and ogt-1 modulate the final level of habituation at a 60s ISI and the rate of habituation but not the final level at a 10s ISI are the first published evidence to support this hypothesis.
It is important to note that the 60s ISI tap habituation phenotype of cmk-1 mutants could be interpreted as a general hyperesponsivity to stimuli throughout the training session, rather than an alteration in habituation – which is the ability of animals to learn to decrement their responding to repeated stimulation (63). However, several lines of evidence from this work suggest that CMK-1 alters habituation. First, when worms were tested at 72 hours post egg lay (Fig. S1; as opposed to 96 hours) cmk-1 mutants displayed wild-type initial responses to mechanical stimuli, but a higher final level of responding at a 60s ISI, dissociating these metrics. Second, cmk-1 mutants display a normal habituated final level when trained at a 10s ISI - a general hyperesponsive phenotype should be present regardless of the stimulus interval used (Fig. 1 A-B). Third, the cmk-1(pg58) mutant displayed larger initial responses, but wild-type habituated responses, genetically dissociating these response metrics (Fig.3B). Taken together, these results suggest that initial and final habituated levels of mechanosensory responding to tap stimuli are independent metrics and that CMK-1 mediates habituation in an ISI-dependent manner.
In conclusion, our results provide the first in vivo evidence of a role for CaMK and O-GlcNAc post-translational modification in responding to mechanosensory stimuli and learning. Additionally, our work provides the first direct evidence in support of the hypothesis that multiple mechanisms mediate habituation at different ISIs and underscores the importance of detailed behavioral genetic analysis to dissect multiple mechanistically independent components of a seemingly simple learned behavior. Finally, our findings will help draw much-needed attention to a common form of posttranslational modification that is required for mechanosensory responding and learning, two fundamental biological processes present in all animals.
Methods
Strains and maintenance
Worms were cultured on Nematode Growth Medium (NGM) seeded with Escherichia coli (OP50) as described previously(64). The following strains were obtained from the Caenorhabditis Genetics Center (University of Minnesota, Minneapolis, MN): N2 Bristol, PY1589 cmk-1(oy21), VC691 ckk-1(ok1033), RB1468 dkf-2(ok1704), VC567 arf-1.2(ok796), VC127 pkc-2(ok328), KG532 kin-2(ce179), RB918 acr-16(ok789), RB818 hum-1(ok634), RB781 pkc-1(ok563), RB1447 chd-3(ok1651), RB830 epac-1(ok655), HA865 grk-2(rt97), NW1700 plx-2(ev773); him-5(e1490), PR678 tax-4(p678), KG744 pde-4(ce268), RB758 hda-4(ok518), RB1625 par-1(ok2001), DA596 snt-1(ad596), XA406 ncs-1(qa406), CB109 unc-16(e109), RB653 ogt-1(ok430), BC10002 dpy-5(e907) and VC40557 (which harbors cmk-1(gk691866) among many other mutations (65)). The following strains were obtained from the National Bioresource Project for the nematode (School of Medicine, Tokyo Womens Medical Hospital, Shinjuku-ku, Japan): FX01046 ogt-1(tm1046), FX01282 T23G5.2(tm1282), FX03075 pdhk-2(tm3075), FX00870 nhr-6(tm870), FX04733 syx-6(tm4733), FX05136 R11A8.7(tm5136), and FX02653 rab-30(tm2653).
Transgenic strains
The transgenic C. elegans strain VH905 hdIs30[Pglr-1::DsRed2] was a gift from H. Hutter (Simon Fraser University, Burnaby, BC). The plasmid containing Pmec-7::mRFP was a gift from J. Rand (University of Oklahoma Health Sciences Center, Oklahoma City, Oklahoma). The transgenic C. elegans strains YT1128 lin-15(n765); tzEx[Pckk-1::GFP; lin−15(+)] and YT2016 tzIs2[Pcmk-1::GFP; rol-6(su1006)] and plasmids containing cmk-1 cDNA were gifts from Y. Kimura (Mitsubishi Kagaku Institute of Life Sciences, Japan). Please see supplemental methods for the primer sequences for PCR fusion constructs generated for this study.
The following strains were created for this work: VG183 yvEx64[Pcmk-1::GFP; Pmec-7::mRFP], VG12 hdIs30[Pglr-1::DsRed2]; tzIs2[Pcmk-1::GFP; rol-6(su1006)], VG19 tzEx[Pckk-1::GFP; lin-15(+)]; hdIs30[Pglr-1::DsRed2], VG92 cmk-1(oy21); yvEx49[Pcmk-1::CMK-1; Pmyo-2::GFP], VG100 cmk-1(oy21); yvEx57[Pcmk-1::CMK-1; Pmyo-2::GFP], VG260 yvEx73[Pogt-1::GFP; Pmec-7::RFP; rol-6(su1006)] VG214 yvEx70[Pogt-1::GFP; rol-6(su1006)] and VG261 yvEx74[Pogt-1::GFP; Pmec-7::RFP; rol-6(su1006)] VG271 cmk-1(oy21); dpy-5(e907), VG279 cmk-1(gk691866); dpy-5(e907), VG245 cmk-1(oy21); ogt-1(ok430).
Imaging procedures
Adult worms were anesthetized in 100 mM NaN3 dissolved in M9 buffer containing sephadex beads (G-150-50, Sigma-Aldrich, St. Louis, MO) on glass microscope slides, and then covered with a 1.5 thick coverslip. An Olympus Fluoview 1000 Confocal microscope was used for imaging. GFP was excited using a 488 nm wavelength laser setting with light emitted collected through a 491-515 nm bandpass filter. dsRed and mRFP were excited using a 543 nm wavelength laser setting with light emitted collected through a 600-630 nm bandpass filter. Optical sections of 0.5 μm thickness were collected using a 60x oil immersion lens (Olympus). Final figures were generated using Image J version 1.41o (National Institutes of Health, Bethesda, MD) and Adobe Photoshop 7.0 (Adobe Systems, San Jose, CA).
Behavioral testing of mutant strains
Worms were synchronized for behavioral testing by picking 5 gravid adults onto a Petri plate containing nematode growth media (NGM) seeded with 50 μl of a liquid culture of OP50 E. coli 12-24 hours earlier and letting them lay eggs for 3-4 hours before they were removed. These eggs were allowed to develop for 96 hours (unless otherwise stated) in a 20°C incubator. Plates of worms were placed into the tapping apparatus (66) and covered with an optically transparent lid constructed from a Petri plate lid, non-fogging cover-glass and wax. After a 100s acclimatization period, 30 taps were administered at either a 60s or a 10s ISI.
For CMK-1 rescue strains twelve hours prior to testing, 40-60 worms carrying the selection marker were transferred using a platinum pick to a fresh NGM plate. Plates were seeded with 50 μl of a liquid culture of OP50 E. coli 16-20 hours beforehand.
Complementation test
Wild-type or cmk-1(oy21) males were mated with dpy-5(e907) hermaphrodites homozygous for one of the three cmk-1 alleles (wild-type, oy21, or gk691866). Tap habituation behavior of non-dpy F1 progeny was evaluated.
Image acquisition of behavior, Behavioral scoring and statistical analysis
Stimulus delivery and image acquisition to record the behavior of the worms was done using the Multi-Worm Tracker (version 1.2.0.2) (66) as described previously (30). Offline data analysis was performed using Choreography analysis software (version 1.3.0_r1035 software package) (66) as described previously (30).
Reversal distances and durations in response to tap were compared across strains by statistical analysis of variance and Tukey honestly significant difference (HSD) tests. Genotype was modeled as a fixed effect. Petri plate (on which the worms were tested; minimum of 3 Petri plates of ~ 50 worms per experimental condition) was modeled as a random effect nested within the fixed effect. For all statistical tests an alpha value of 0.05 was used to determine significance. All analysis was performed using the statistical software programming language, R, and the following R packages: tidyverse, stringr, nlme, broom and multcomp.
Analysis code can be accessed here: https://github.com/ttimbers/CaMK_and_OGT-1_modulate_mechanoresponding_and_learning/tree/master/src
Kinase and phosphosite prediction and evolutionary analyses
The kinase substrate specificity prediction matrices (KSSPM) for C. elegans CMK-1, human CaMK1 isoforms and human CaMK4 were generated using an updated version of the algorithm originally described in Safaei et al. (33). The C. elegans CMK-1 KSSPM was used to score all of the hypothetical peptides predicted surrounding each of the serine and threonine residues in the 20,470 known C. elegans protein sequences. The top 597 scoring phosphopeptides were examined for their conservation in humans using the algorithm described in Safaei et al. (33). The identified human phosphosites were then scored with the KSSPMs for human CaMK1 isoforms and human CaMK4.
Acknowledgements
We would like to thank Angela Leong, Jing Xu and Savannah Nijeboer for help running experiments and Andrew C. Giles for useful advice and discussions regarding this research. This work was supported by a Natural Sciences and Engineering Research Council Alexander Graham Bell Canada Graduate Scholarship to TAT, and by a Natural Sciences and Engineering Research Council Alexander Graham Bell Canada Graduate Scholarship to ELA, and by a Natural Sciences and Engineering Research Council Discovery Grant to CHR.
References




