

Abstract
Zika virus (ZIKV) is a flavivirus that has recently been associated with increased incidence of neonatal microcephaly and other neurological disorders. The virus is primarily transmitted by mosquito bite, although other routes of infection have been implicated in some cases. The Aedes aegypti mosquito is considered to be the main vector to humans worldwide, but there is evidence of other mosquito species, including Culex quinquefasciatus, playing a role in the Brazilian outbreak. To test this hypothesis, we experimentally compared the vectorial competence of laboratory-reared A. aegypti and C. quinquefasciatus. We found ZIKV in the midgut, salivary glands, and saliva of artificially fed C. quinquefasciatus. Additionally, we collected ZIKV-infected C. quinquefasciatus from urban areas of high microcephaly incidence in Recife, Brazil. Take into account; these findings indicate that there may be a wider range of vectors for ZIKV than anticipated.
Zika is classically considered a mild disease whose symptoms include fever, joint pain, rash and, in some cases, conjunctivitis (1). However, the Zika outbreak in Brazil has been associated with an increased incidence of neonatal microcephaly and neurological disorders (2, 3). Zika virus (ZIKV) is a poorly known, small, enveloped RNA virus with ssRNA (+) belonging to the Family Flaviviridae. It was first isolated in April 1947 from a rhesus monkey and in January 1948 from the mosquito species Aedes africanus (4). Since then, several ZIKV strains have been isolated from many samples, mostly mosquitoes, including species from the genera Aedes, Mansonia, Anopheles and Culex (5).
The first known Zika epidemic in an urban environment occurred in Micronesia in 2007, with approximately 73% of the human population on Yap island becoming infected (6). Intriguingly, although many Aedes mosquitoes were collected in the field and evaluated for virus detection, no samples were found to be positive for ZIKV (6). Additionally, it is important to highlight that Aedes aegypti (A. aegypti) is absent from most islands in the Micronesia archipelago and is very rare on the islands where it is present (6, 7).
There is a global consensus among scientists and health agencies that Aedes spp. are the main ZIKV vector in urban areas (WHO, 2016). This is in part because vector competence experiments for ZIKV have been conducted exclusively for species of this genus, mainly A. aegypti (8, 9). Previous laboratory studies (8, 10) suggested that A. aegypti is a ZIKV vector. Recently, high rates of dissemination and transmission of the ZIKV in A. aegypti has been observed under laboratory conditions (11). Intriguingly, a few studies show that A. aegypti and Aedes albopictus populations have low rates of ZIKV transmission (12) or none (13, 14), but the role of other vectors in the spread of ZIKV has been overlooked. Thus, other mosquito species, co-existing with A. aegypti in urban areas, could contribute to ZIKV transmission (15). Here, we report data that support the idea that Culex quinquefasciatus, the most common mosquito in tropical and subtropical areas, is a potential ZIKV vector. We performed mosquito vector competence assays under laboratory conditions, comparing both A. aegypti and C. quinquefasciatus using different virus doses, as well as the detection of ZIKV in wild C. quinquefasciatus mosquitoes. ZIKV was detected in salivary glands and in the saliva of artificially fed C. quinquefasciatus mosquitoes, suggesting that this species is a potential vector for ZIKV transmission. Taken together, our results have implications for vector control strategies and understanding the epidemiology of ZIKV.
Vector competence assays
Artificial blood feeding assays were performed using two laboratory-reared colonies: RecLab (A. aegypti) and CqSLab (C. quinquefasciatus) and a field-collected population of A. aegypti(F1/F2) from the Archipelago of Fernando de Noronha, a district from Pernambuco state, Northeast Brazil. A local Zika virus strain, isolated from the serum of a patient with Zika symptoms from Pernambuco State, Brazil, during the 2015 outbreak (ZIKV/H.sapiens/Brazil/PE243/201), fully characterized (accession number KX197192.1), was used in vector competence assays.
Seven to ten days-old females were challenged in an artificial feeding, consisted of a Petri dish covered with Parafilm M®, with a mixture containing equal volumes of defibrinated rabbit blood and the viral suspension. Here, in each assay, we used two different viral doses: 106 PFU/ml and 104 PFU/ml. Mosquitoes were exposed for 90 minutes and, after that, only the engorged females were transferred to another cage and maintained in the infectory room, under bio safety conditions (BSL2) for 15 days. At 3, 7, 11 and 15 days post infection (dpi), midguts and salivary glands were dissected individually and transferred to 1.5 ml vials containing a mosquito diluent (20% of fetal bovine serum in PBS with 50 μg/ml penicillin/streptomycin, 50 μg/ml gentamicin and 2.5 μg/ml fungizone) and stored at −80°C until further usage. After RNA extraction, samples were assayed by quantitative RT-PCR (RT-qPCR) and both Infection Rate (IR), which is the proportion of infected midguts, and the proportion of infected salivary glands (SR), which is the number of positive salivary glands divided by the total number of salivary glands tested, were calculated for each species on each dpi. All procedures are described in details in the Supplemental Materials.
A total of 289 mosquitoes were examined for ZIKV infection by RT-qPCR. Among these mosquitoes, 130 were A. aegypti RecLab, 60 were A. aegypti FN and 99 were C. quinquefasciatus. During the extrinsic incubation period, a high mortality rate was observed in the infected group a few days after blood feeding, with peak mortality observed between 3-5 dpi. In A. aegypti populations, the mortality rate ranged from 48% to 52%; in C. quinquefasciatus mosquitoes, it ranged from 33% to 44% (data not shown).
In both species, we detected ZIKV in the midgut at most time-points under study, except in field-collected A. aegypti (FN) blood fed at a low viral dose (104 PFU/ml). In the salivary glands of A. aegypti, we detected ZIKV-positive samples at 3 dpi for both viral doses. Although variations in infection were observed, the IR between Aedes and Culex mosquito’s species was not statistically significant (Fig 1A). Analysing only Culex species, the differences in IR were not statistically significant comparing different viral doses (p>0.05). When a high viral dose (106 PFU/ml) was used during artificial feeding, the SR reached an average of 60% in A. aegypti RecLab, and 100% in C. quinquefasciatus at 7 dpi, but this average declined to 20% at 15 dpi in Culex and to 50% in A. aegypti. However, this difference was not significant (p>0.05) (Fig 1B). When a lower viral dose was used, for both A. aegypti populations (lab and field-caught), we observed that the mosquito lab colony infection rate was higher than field-caught mosquitoes (Figure 1A p=0.0186), although there were no differences in SR (Figure 1B p>0.05). The maximum SR in Culex at lower viral dose, was 10% at 3dpi declining to zero after 15 dpi (p>0.05) (Fig. 1B). We also sampled mosquitoes at 11 dpi in the first trial, except for field-collected A. aegypti (FN), which had a SR of 10%; neither C. quinquefasciatus nor A. aegypti samples from the lab were ZIKV positive (data no shown).
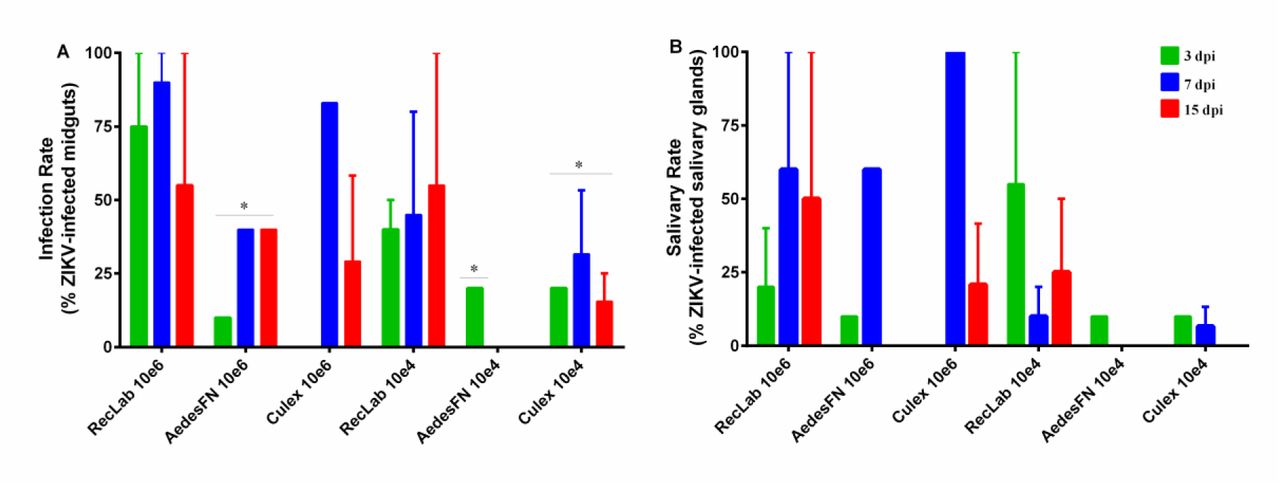
Experimental infection of ZIKV in laboratory-reared A. aegypti and C. quinquefasciatus collected 3 (green bars), 7 (blue bars) and 15 days post infection (dpi) (red bars). (A) Proportions of ZIKV-positive midguts at each sampling point (average mosquitoes per group = 10 for two replicates). (B) Proportions of ZIKV-positive salivary glands at each sampling point (average of mosquitoes per group = 10 for two replicates). Significance was determined using one-way ANOVA with Tukey’s multiple comparison test (* p < 0.05).
RT-qPCR was used to quantify ZIKV RNA load at the different time-points. In general, viral RNA copies in A. aegypti RecLab in the midguts and salivary glands varied considerably. Both A. aegypti FN and C. quinquefasciatus viral copies in target organs (midgut and salivary glands) remained detectable (Fig. 2A to D). To evaluate ZIKV transmission in saliva for both species, honey-soaked filter papers (FTA Classic Cards, Whatman®, Maidstone, UK) were offered to mosquitoes to feed upon 8-14 dpi. At 9-12 dpi, ZIKV RNA was detected in saliva of both A. aegypti and C. quinquefasciatus species (Fig. 3). When a high viral dose (106) was used, the amount of viral RNA copies expectorated during salivation in both Aedes and Culex were similar at all time-points analysed (p>0.05). However, when the mosquitoes were challenged with a low viral dose (104), A. aegypti expectorated more RNA viral copies than Culex (p=0.0473).

Quantification of RNA viral copy number in the midguts and salivary glands of A. aegypti and C. quinquefasciatus mosquitoes experimentally fed with blood containing ZIKV at 106 PFU/ml (A, B) and 104 PFU/ml (C, D). Squares represent A. aegypti(RecLab) population, inverted triangles represent A. aegypti(FN) population and circles represent C. quinquefasciatus. Significance is shown in the bars and was determined using an unpaired t-test (* p < 0.05, ** p < 0.01).

Quantification of ZIKV in A. aegypti and C. quinquefasciatus saliva expectorated onto FTA cards 9 - 12 days post infection (dpi). Green bars show A. aegypti(RecLab) population blood-fed with ZIKV at 106 PFU/ml, solid blue bars show C. quinquefasciatus population blood-fed with ZIKV at 106 PFU/ml, red bars show A. aegypti(RecLab) population blood-fed with ZIKV at 104 PFU/ml and open blue bars show C. quinquefasciatus population blood-fed with ZIKV at 104 PFU/ml. Significance was determined by an unpaired t-test (* p < 0.05).
Transmission Electron Microscopy
To further confirm our results from RT-qPCR, we performed a transmission electron microscopy from dissected salivary glands from C. quinquesfaciatus infected mosquitoes. The morphological organization of C. quinquefasciatus salivary glands showed an electron-dense apical cavity, displaying membrane projections extending from the wall (Fig. 4A, B). ZIKV infected salivary acinar cells of C. quinquefasciatus showed signs of cytopathic disruptions, including cisternae in the endoplasmic reticulum and tubular proliferated membranes, organized in several patches within a single cell (Fig. 4C, D). Mature ZIKV particles of 40-50 nm in diameter, composed of a central electrodense core (∼30 nm in diameter) surrounded by a viral envelope, were observed inside the dilated endoplasmic reticulum (Fig. 5A to D). In some regions, viral envelope formation is shown to arise from endoplasmic membrane (Fig. 5B). Some ZIKV virions were observed proximal to the apical cavity of the salivary cell. Mitochondria also showed severe damage, including complete loss of cristae (Fig. 5D). In summary, transmission electron microscopy analysis confirmed that C. quinquefasciatus mosquitoes are permissive to ZIKV infection, since viral particles were detected at the salivary glands of artificially fed mosquitoes.

(A-B) Ultrathin sections of uninfected C. quinquefasciatus salivary gland. (A) Shows the electrodense content of the apical cavity (AC) with membrane projections extended from the wall. (B) Uninfected acinar salivary gland cell showing Nu, nucleus; Cyt, cytoplasm; ER, endoplasmic reticulum; Mi, mitochondria. (C-D) Cytopathic effects of salivary glands cells infected with ZIKV showing several patches of tubular proliferated membrane (TPM), distended endoplasmic reticulum (dER) and a phagolysozome-like vacuole (PhV). Cyt, cell cytoplasm; TC; thread-like centers.

Mature ZIKV particles inside C. quinquefasciatus salivary gland cell. (A) Numerous ZIKV within dilated endoplasmic reticulum (dER). (B) Envelope formation from endoplasmic membrane (white arrow). (C) Showing enveloped virus particles with electrodense cores. (D) Viral particles accumulated proximal the acinar cavity (arrows), note damaged mitochondria. Cyt, cell cytoplasm; AC, acinar cavity, Mi, mitochondrion; Vi, virion (s).
ZIKV detection in field-caught C. quinquefasciatus
Lastly, we conducted ZIKV surveillance (February to May 2016) with mosquitoes collected with a battery-operated aspirator device (Horst®) from residences inhabited by individuals with clinical symptoms of zika fever. Field-collected mosquitoes were sorted by place of collection, species, sex, feeding status (engorged and not engorged) and grouped in pools of up to 10 mosquitoes. A total of 1,496 adult C. quinquefasciatus and 408 A. aegypti female mosquitoes were collected from different sites in the Metropolitan Region of Recife (Fig. S1). These mosquito pools were grinded in Leibovitz medium supplemented with 5% FBS. These samples were separated into two aliquots, one for RT-qPCR and the other for virus isolation. From 270 pooled-samples of adult female C. quinquefasciatus and 117 pools of A. aegypti mosquitoes assayed by RT-qPCR, three Culex and two Aedes pools were positive for ZIKV. Interestingly, two out of the three positive Culex samples were not blood-fed, whereas concerning Aedes pools, the two positive pools for ZIKV were fed. The cycle threshold (Ct) of Culex positive pools when screened by RT-qPCR were 37.6 (sample 5), 38.0 (sample 17) and 38.15 (sample 163). Concerning Aedes pools, Cts were 37.5 (sample 3) and 37.9 (sample 7). Minimum infection rate (MIR - number of positive pools divided by the total of specimens assayed multiplied by 1000) were calculated for both species. For C. quinquefasciatus, MIR was 2.0 and concerning A. aegypti, MIR was 4.9. In an attempt to isolate ZIKV from field-caught Culex mosquitoes, we inoculated in African Green Monkey kidney cells, samples from two positive pools with the lowest cycle thresholds. Indeed, ZIKV was isolated from these samples, thus unambiguously demonstrating that this species was carrying active ZIKV particles in Recife, Brazil. Two ZIKV-positive isolates from field-caught Culex mosquitoes were submitted to Sanger and MinION platforms. Sanger sequencing resulted in low quality sequences and only a partial fragment was acquired from MinION sequencing, probably as a direct result of low viral titers. This partial sequence enabled us to confirm the virus identity. Sequence was deposited at GenBank, and accession number is still to be provided.
Discussion
Our work has associated a second mosquito genus in ZIKV transmission cycle in Northeastern Brazil. We showed that, C. quinquefasciatus, also known as the southern house mosquito, which is the most common mosquito in urban areas in Brazil, is susceptible to infection with ZIKV during experimental blood feeding; moreover, we found that ZIKV has an active replication cycle in the salivary glands and being subsequently released in the saliva. In addition, we were able to detect ZIKV circulating in wild C. quinquefasciatus collected from Recife.
Although it is widely assumed that A. aegypti is the main ZIKV vector, previous vector competence studies are inconclusive. In the present study, a low dose of ZIKV (104 PFU/ml) was used for comparison with the higher doses used in previous studies (11, 12, 14). We found that both A. aegypti and C. quinquefasciatus can be experimentally infected by ZIKV even at low doses and that ZIKV virus was subsequently detected in the saliva.
Cornet et al. (10) concluded that not all infected mosquitoes could transmit the virus and could not always transmit it, in contrast to the idea that once infected, a mosquito would transmit virus for its entire life. This finding suggests that a time window for vector-borne ZIKV transmission may exist. We found that after 11 dpi, most samples were negative for ZIKV (apart from one positive salivary gland of A. aegypti given 106 PFU/ml), thus our maximum time point analysis was set to 15 days post infection. However, Boorman & Porterfield (8) reported that virus replication resumed at 15-20 dpi and ZIKV remained present in Aedes mosquitoes for up to 60 days.
To confirm that the virus detected in the salivary glands by RT-qPCR was being released in saliva during consecutive blood meals, we followed up the viral load from days 8 to 14 post-infection using filter paper cards. This strategy of viral RNA detection directly from FTA cards has been employed in previous studies for arbovirus surveillance (16, 17). In the present study, we successfully detected ZIKV RNA copies in cards from A. aegypti and C. quinquefasciatus populations. This result demonstrates that in addition to being susceptible to ZIKV infection, allowing virus replication in the salivary glands, both species are capable of effectively transmit ZIKV.
RT-qPCR results were confirmed by transmission electron microscopy. The general mature ZIKV morphology observed on the salivary glands confirmed previous ultrastructural studies (18-20). In salivary glands cells, ZIKV replication causes cytopathic effects by 7 dpi. Similar results have been shown for West Nile virus (WNV) (21, 22), although we did not directly observe ZIKV nucleocapsids budding from endoplasmic reticulum membranes nor from the tubular proliferated membrane (21). The fact that we found salivary glands positive for ZIKV when the midgut of the same mosquito was negative indicates that mosquitoes may be clearing viral infection in the midgut while virus replication continues in the salivary glands. This finding has implications for the analytical methods employed in vector competence studies.
Currently, there is a lack of studies investigating Culex vectorial competence for ZIKV. Most studies have targeted only Aedes species, and only a few studies have compared different species (including Culex) regarding natural infection rates. Surprisingly, Diallo et al. (5) observed a higher minimum infection rate for Culex perfuscus (10x higher) than for A. aegypti. Positive A. aegypti samples have always been reported at very low infection rates, even in areas with high human ZIKV infection rates, such as Malaysia (23). Indeed, in Micronesia (6), and French Polynesia, ZIKV was not detected in wild-caught Aedes spp. mosquitoes during outbreaks. It is interesting that in all of these areas, C. quinquefasciatus is an abundant mosquito species that may have also played an undetected role in ZIKV transmission. Furthermore, A. aegypti and C. quinquefasciatus have completely different behaviours regarding feeding periods and breeding site preferences.
Thus, our findings indicate that vector control strategies may need to be re-examined since reducing A. aegypti populations may not lead to an overall reduction in ZIKV transmission if Culex populations are slight affected by Aedes specific control measures. To the moment, there is no broad ongoing program for C. quinquefasciatus control in Brazil, although Recife, Olinda and Jaboatão dos Guararapes, three municipalities in Recife Metropolitan Region, have undertaken specific control of C. quinquefasciatus to control lymphatic filariasis transmission locally (24).
Viral transmission via C. quinquefasciatus is not a new concept; this species is the major vector of West Nile virus in North America (25), along with Japanese encephalitis virus (26) and equine encephalitis virus (27). Our present study indicates that C. quinquefasciatus mosquitoes may be involved in ZIKV transmission in Recife. Thus, it is now necessary to understand the contributions of each species in transmission to target each one properly. In conclusion, considering its high abundance in urban environments and its anthropophilic behaviour in Brazil (28-30), C. quinquefasciatus may be a vector for ZIKV in this region.
Acknowledgements
This work was supported in part by the Fundação de Amparo à Pesquisa do Estado de Pernambuco (FACEPE; APQ-1608-2.13/15 and APQ-0085-2.13/16 to C.F.J.A.) and the National Institute of Allergy and Infectious Diseases of the National Institutes of Health (R01AI095514 and 1R21AI128931-01 to W.S.L.) C.F.J.A. and C.A.P. are supported by productivity fellowship from the Brazilian National Council for Research and Development (CNPq). We thank the staff of the insectary at Aggeu Magalhães Research Center for technical assistance, the Program for Technological Development in Tools for Health (PDTIS-FIOCRUZ) for allowing us to use their facilities, and the staff of the Pernambuco State Health Department for sharing recent data on microcephaly and assisting in surveillance.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}