

Abstract
Summary Most plant-feeding insects are ecological specialists restricted to one or a few closely related host-plant species (Forister et al. 2015). A long-standing hypothesis asserts that natural selection favors host specialization because trade-offs between performance on alternative host species limit the fitness of generalists, yet empirical evidence for such trade-offs is scarce (Futuyma and Moreno 1988; Forister et al. 2012). Here we show that trade-offs between adaptations to alternative hosts occur over both long‐ and short-term macroevolutionary timescales, but positive associations between host-use traits are also abundant. Host-use records of 1604 caterpillar (Lepidoptera) species revealed negative associations between adaptations to two diverse groups of host-plant taxa over 150 million years (Misof et al. 2014) of caterpillar evolutionary history, but a different division between use of angiosperm and pine hosts among closely related caterpillars. In contrast, host-use records of 955 true bug (Hemiptera) species suggested uniformly positive associations between adaptations to the same host taxa both over the 300-million-year (Misof et al. 2014) evolutionary history of true bugs and among closely related species. The lack of consistent patterns across insect orders and timescales suggests that host-use trade-offs are historically contingent rather than universal constraints, reflecting the diversity of mechanisms driving host-specialization in plant-feeding insects.
Main Text
The ubiquity of ecological specialists (Forister et al. 2012, 2015) is often attributed to evolutionary constraints on generalism (Futuyma and Moreno 1988; Futuyma et al. 1995). Adaptations to one environment may come at the cost of performance in other environments (Futuyma and Moreno 1988; Forister et al. 2012), and such trade-offs are crucial elements of most theoretical models of the evolution of specialization (Ravigné et al. 2009). In many biological systems, trade-offs are implied by negative correlations between traits (Shoval et al. 2012), but performance measures on alternative host species are most often uncorrelated or positively correlated in wild and experimentally selected populations of plant-feeding insects (Futuyma 2008; Forister et al. 2012). In fact, although plant-feeding insects are a model system for the study of ecological specialization (Futuyma and Moreno 1988; Forister et al. 2012), the role of trade-offs in the evolution of insect host-specialization remains generally unsupported.
However, trade-offs that drive specialization in plant-feeding insects may be difficult to detect within species (Joshi and Thompson 1995). Host-use traits are phylogenetically conserved in many insect groups (Futuyma and Agrawal 2009), and genetic variation for use of novel hosts is often absent within a single population (Futuyma et al. 1995). These constraints likely stem from the fact that resource use is a complex trait involving multiple interacting genetic loci (Remold 2012), so the evolutionary trajectories that lead to high fitness on alternative resources may be mutually exclusive. For example, the evolution of improved performance on one host can select for a preference for that host, driving a positive feedback loop of further specialization (Fry 1996). When epistatic interactions are strong, the greatest variation in host-use strategies will evolve between reproductively isolated lineages over long time scales (Satterwhite and Cooper 2015), with trade-off outcomes determined by each lineage’s evolutionary history (Rodriguez-Verdugo et al. 2014). Analagous macroevolutionary trade-offs have been described in plants; the presence of alternative defensive strategies are negatively correlated over plant evolutionary history (Campbell and Kessler 2013). However, it remains unknown whether this functional diversification across hosts has created macroevolutionary trade-offs for plant-feeding insects; performance on alternative hosts may instead be positively associated if generalist adaptations have been prevalent during the evolution of insect herbivores (Peterson et al. 2015).
We investigated trade-offs between adaptations to diverse host taxa over long‐ and short-term macroevolutionary timescales in two orders of plant-feeding insects: Lepidoptera (caterpillars) and Hemiptera (true bugs). Assuming that trade-offs should produce negative correlations between recorded presence on alternative host taxa for individual insect species (Peterson et al. 2015), we quantified the macroevolutionary relationships between use of the most common host-plant orders for North American insects (Figure 1). Long‐ and short-term effects were partitioned by estimating separate correlation parameters for phylogenetic and residual effects (Figure 2) in a phylogenetic mixed model. We assembled the pairwise correlations between use of all focal hosts to produce network graphs of host-use relationships, revealing overall patterns of host-use evolution in each insect order. Specifically, we investigated two potential trade-off scenarios: segregation of hosts into discrete clusters and isolation of individual host orders.
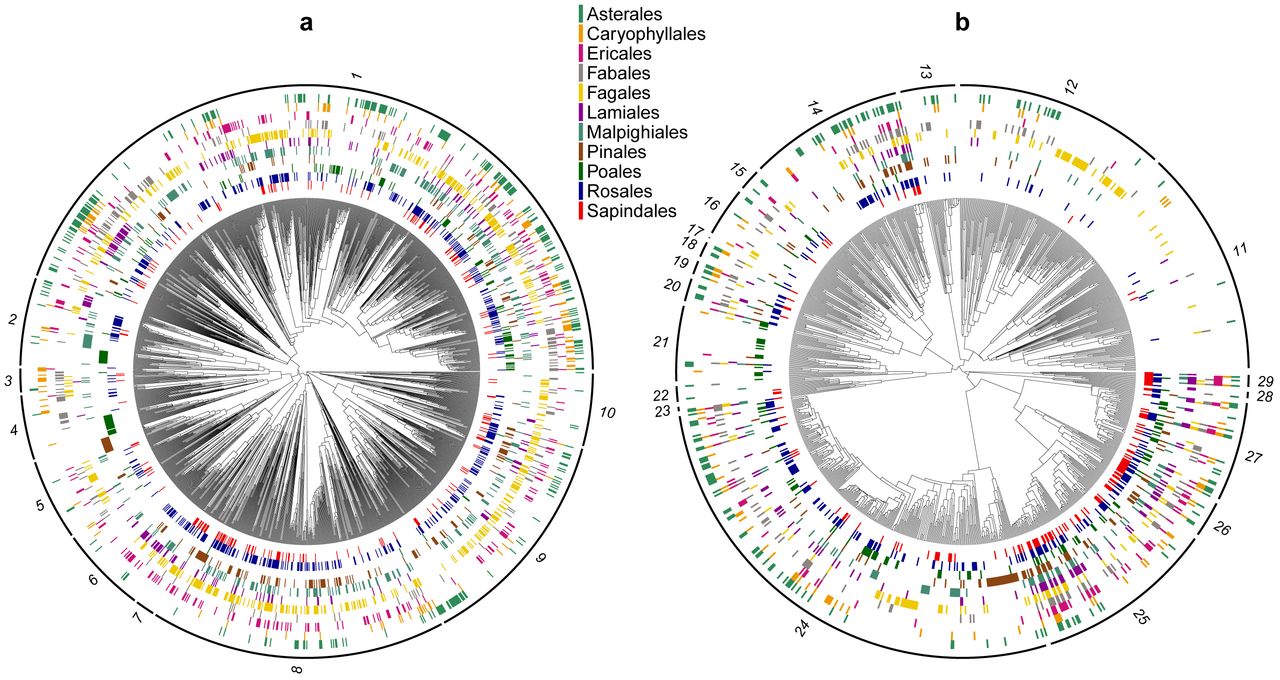
For each host-plant order, colored blocks indicate which insect species have been observed on that host. Insect species with no hosts shown were observed only on non-focal hosts or had no host-use information associated with their locality records (Hemiptera only). Insect families are indicated around the phylogenies as follows: a, Lepidoptera – 1: Noctuoidea, 2: Nymphalidae, 3: Lycaenidae, 4: Hesperiidae, 5: Pyralidae, 6: Sphingidae, 7: Saturniidae, 8: Geometridae, 9: Tortricidae, 10: Gracillariidae. b, Hemiptera – 11: Cicadellidae, 12: Membracidae, 13: Cicadidae, 14: Miridae, 15: Tingidae, 16: Pentatomidae, 17: Scutelleridae, 18: Coreidae, 19: Rhopalidae, 20: Lygaeidae, 21: Delphacidae, 22: Fulgoridae, 23: Flatidae, 24: Aphididae, 25: Diaspididae, 26: Coccidae, 27: Pseudococcidae, 28: Psylloidea, 29: Aleyrodidae.

a, Negative phylogenetic correlation. b, Positive phylogenetic correlation. c, Negative residual correlation. d, Positive residual correlation. e, Negative phylogenetic and positive residual correlations. f, Positive phylogenetic and negative residual correlations. In each example, filled red squares on the left indicate which species in the phylogeny are present on host 1, and filled blue squares on the right indicate which species are present on host 2.
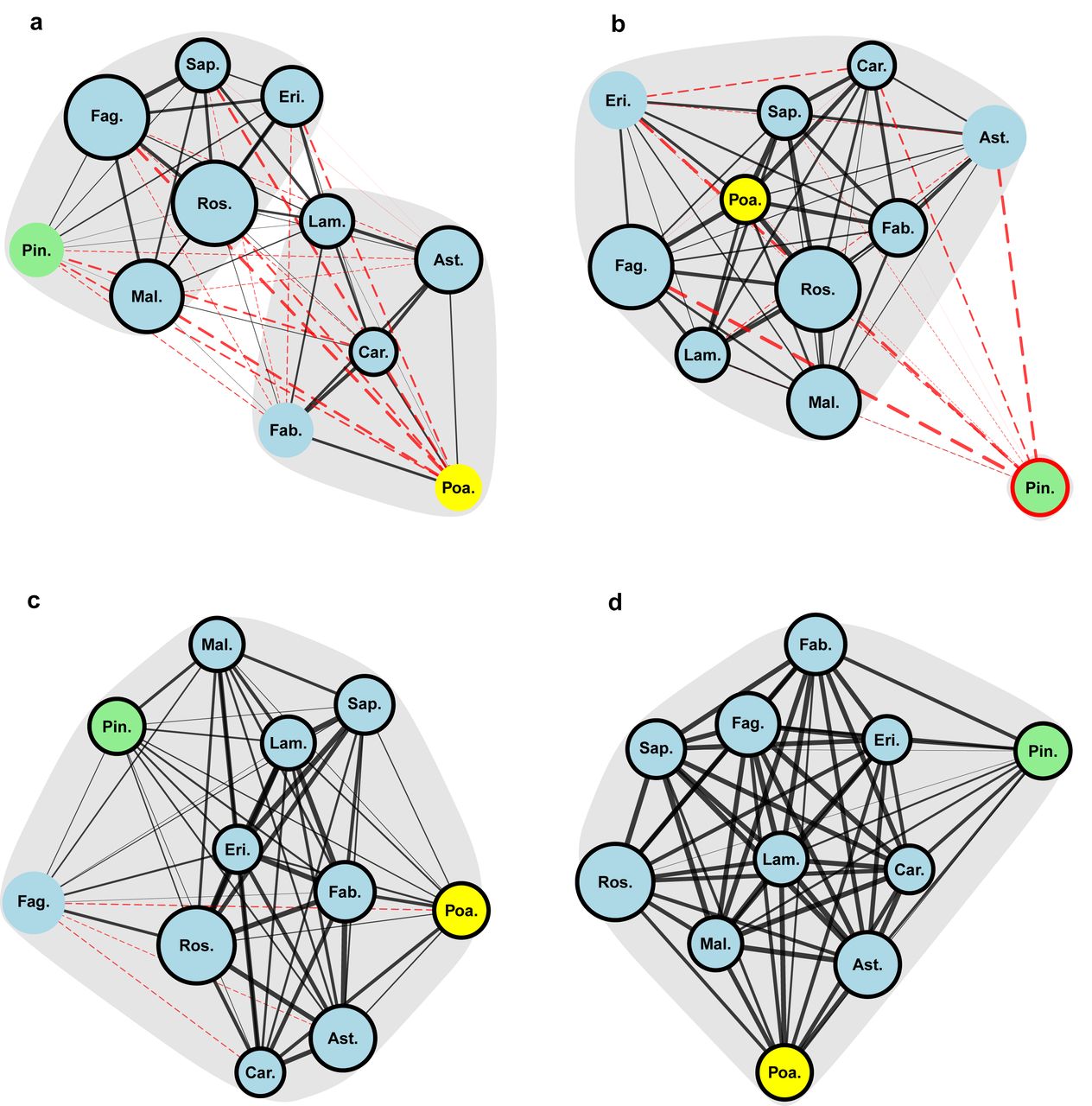
We found that the network of long-term evolutionary relationships between lepidopteran use of the focal host orders was significantly structured (P < 0.01), revealing two large clusters of host taxa (Figure 3a). Clustering is a common feature of ecological networks (Olesen et al. 2007) that implies functional relationships between organisms which often correlate with phylogenetic distances (Krasnov et al. 2012; Hadfield et al. 2014). However, cluster membership in our results was surprising: the gymnosperm Pinales (pines) and monocot Poales (grasses) were each affiliated with a different set of eudicot orders. Instead of host-insect coevolution, long-term evolutionary trade-offs experienced by lepidopteran lineages appear to reflect a broad-scale ecological division between plant hosts. The relevant ecological traits that differ between the two host clusters are not obvious, but may correspond to the habitat types where those hosts are found (Futuyma 1976).

a, Lepidoptera – phylogenetic correlations. b, Lepidoptera – residual correlations. c, Hemiptera – phylogenetic correlations. d, Hemiptera – residual correlations. Each vertex represents a host order, with vertex area proportional to the number of insects that were observed on that host. Positive interactions between presence a pair of hosts are represented by solid, black lines and negative correlations by dashed, red lines, with line thickness proportional to the magnitude of the correlation. Network spatial structure was determined using the Kamada-Kawai algorithm (Kamada and Kawai 1989), a force-directed layout method in which “repulsion” between vertices was proportional to the inverse of one plus the correlation values between the respective hosts. Vertices are labelled with the following abbreviations – Ast: Asterales, Car: Caryophyllales, Eri: Ericales, Fab: Fabales, Fag: Fagales, Lam: Lamiales, Mal: Malpighiales, Pin: Pinales, Poa: Poales, Ros: Rosales, Sap: Sapindales. Vertices are colored by taxonomic group – Eudicots: blue, Monocots: yellow, Conifers: green. Statistically significant modules (P < 0.05) are indicated by grey bubbles. Individual host orders with mean correlations of significantly higher magnitude than expected (P < 0.05) are indicated by bold vertex outlines (black for positive means, red for negative means).
Short-term correlations between lepidopteran use of the focal host taxa also showed significant network structure (P < 0.01) but use of all angiosperm hosts formed a single cluster of mostly positive associations (Figure 3b). The disappearance over short evolutionary timescales of the trade-off between angiosperm hosts suggests that individual generalist species have overcome the constraints of historical contingency rather than merely maximizing host breadth within their lineage’s niche. On the other hand, we do not account for differences in fecundity between specialist and generalist insects on particular hosts; it may be that generalists usually have lower fitness – i.e. they are jacks of all trades but masters of none (Futuyma and Moreno 1988). However, costs of generalism have been difficult to document (Forister et al. 2012; Gompert et al. 2015), so the positive residual correlations we observed may instead represent evolutionary breakthroughs made possible by novel mechanisms of phenotypic plasticity or other generalist adaptations (Barrett and Heil 2012).
Nevertheless, although we observed few constraints to short-term evolution of lepidopteran host-use within the focal angiosperm taxa, we did find evidence for a short-term evolutionary trade-off between use of angiosperms and Pinales (pines). Lepidopteran presence on pines and many of the focal angiosperm taxa were positively correlated over long timescales, yet individual lepidopteran species found on pines were less likely to use all other focal host taxa (mean pairwise correlation -0.23, P < 0.01). This pattern suggests that although pines and some angiosperms are used by insects with similar evolutionary backgrounds, a trade-off between evolutionarily labile traits tends to prevent the simultaneous use of those host taxa by individual caterpillar species. Such trade-offs between labile but mutually exclusive host-use traits may be particularly significant because they can promote rapid speciation (Nosil et al. 2002), driving the adaptive radiations that have produced much of the plant-feeding insect diversity on earth (Farrell 1998; Janz et al. 2006).
In contrast to the observed patterns in the Lepidoptera, hemipteran host-use showed no evidence of trade-offs over long or short evolutionary timescales. Associations between hemipteran presence on all focal host orders were uniformly positive (Figure 3c,d) and network structure showed significant support for a single host-use cluster encompassing all focal hosts for both phylogenetic (P < 0.01) and residual correlations (P < 0.01). These results suggest that generalist adaptations that increase fitness across multiple hosts have been more important for Hemiptera than specialist adaptations to particular hosts (Peterson et al. 2015). Moreover, hemipteran generalism appears completely unrestrained by host taxonomy even over long time scales, leading to the evolution of both super-generalist species and clades (Normark and Johnson 2011). The trade-offs observed in lepidopteran host-use are therefore not universal to plant-feeding insects; they depend on evolutionary context.
There are many differences between Lepidoptera and Hemiptera, but their fundamentally distinct relationships with host plants may be particularly important to understanding why the role of trade-offs appears to be very different in the two groups. Hemiptera are sucking insects, while Lepidoptera are generally leaf-chewers (Forister et al. 2015). These two feeding modes elicit different modes of plant defensive responses (Ali and Agrawal 2012), and sap-sucking may be particularly amenable to generalist adaptations that circumvent host defenses (Barrett and Heil 2012). In contrast, Lepidoptera often rely on specialized enzymes to detoxify defensive chemicals, which may constrain the evolution of generalism, although super-generalist Lepidoptera do exist, possibly powered by phenotypic plasticity in enzyme expression (Li et al. 2002).
Overall, we found strong evidence that both labile traits and long-term, historically contingent evolutionary processes produce important host-use constraints in plant-feeding insects. Nevertheless, the relatively few, broad-scale trade-offs found here fail to explain the observation that a vast majority of plant-feeding insects are restricted to a single host family (Forister et al. 2015). It is clear that tradeoffs can vary based on genetic context (Rodriguez-Verdugo et al. 2014), so we cannot rule out the hypothesis that species-specific, idiosyncratic trade-offs drive ecological specialization. Nevertheless, although individual species-specific trade-offs have been documented (Nosil et al. 2002), our results suggest that most microevolutionary trade-offs do not persist over macroevolutionary timescales. Trade-offs play an intuitive and possibly inescapable role in constraining performance across multiple tasks (Shoval et al. 2012), yet host-specialization in plant-feeding insects is not necessarily driven by host-specific limitations on survival and growth. Alternative factors, such as neural constraints (Bernays 2001), mate-finding (Hawthorne and Via 2002), or genetic drift (Gompert et al. 2015) may ultimately shape the evolution of each species’ ecological niche.
Supplementary Information is available in the online version of the paper.
Author Contributions
All authors were involved in the study design and wrote the manuscript. D.A.P. and N.B.H. analysed the data.
Author Information
The authors declare no competing financial interests. Correspondence and requests for materials should be addressed to D.A.P. (dapeters{at}cns.umass.edu)
Materials and Methods
Data
Lepidopteran host-use data were downloaded from the HOSTS database (nhm.ac.uk/hosts) (Robinson et al. 2015), a collection of published records of worldwide caterpillar host-plants. Hemipteran host-use data were downloaded from the Tri-Trophic Thematic Collection Network database (tcn.amnh.org), a compilation of insect collection records from academic and governmental museums in the United States. For both datasets, we restricted our analysis to records from North America (all localities labelled USA, Canada, Mexico or Nearctic). All plant taxonomic names were standardized with the Taxonomic Name Resolution Service (Boyle et al. 2013) and insect taxonomic names with the python package TaxonNamesResolver and the following reference databases: Aphid Species File (Favret 2015), Integrated Taxonomic Information System (itis.gov), and Catalogue of Life (catalogueoflife.org).
We created binary presence/absence matrices of lepidopteran and hemipteran species by host plant taxon, with insects considered present on all hosts for which they had at least one host-use record. To focus computational resources on host taxa with enough statistical power to detect evolutionary host-use interactions, we restricted our analyses to focal host orders used by at least 100 insect species in one insect order (~10% of the total focal insect species per order). Eleven host orders met this cutoff, and each of these met the cut-off for both Hemiptera and Lepidoptera: Asterales, Caryophyllales, Ericales, Fabales, Fagales, Lamiales, Malpighiales, Pinales, Poales, Rosales, and Sapindales. Interactions with these focal orders accounted for 77% of total interactions in the Lepidoptera dataset and 57% of total interactions in the Hemiptera dataset.
We characterized shared ancestry among the North American lepidopteran and hemipteran species in our host-use dataset by estimating time-scaled phylogenies using a phyloinformatic approach (see Supplemental Materials for details). Phylogenetic data were not available for all species in the host-use dataset, but there was an overlap of host-use and phylogenetic data for 1604 lepidopteran species and 955 hemipteran species. Phylogenies and host-use matrices for these species will be available on Dryad (datadryad.org) upon publication.
Statistical Analysis
We used a phylogenetic meta-analysis approach to test for evolutionary correlations between insect use of the focal host-plant orders (Hadfield and Nakagawa 2010). Treating each pairwise comparison between host taxa individually, we set up a phylogenetic mixed model for each insect order using the package MCMCglmm (Hadfield 2010) in the R statistical framework (R Core Team 2015). We estimated the evolutionary interaction between the pair of host traits using the “random=~us(trait):Insect” and “rcov=~us(trait):units” syntax to capture both phylogenetic and residual effects. Prior parameter distributions were specified as “prior<-list(R=list(V=diag(2),nu=2), G=list(G1=list(V=diag(2),nu=2)))”. All chains were run for 10 million iterations with a burn-in of 1 million iterations, and we ran ten MCMC chains for each model to test for convergence. Gelman-Rubin convergence analysis of the ten chains for each model produced potential scale reduction factors under 1.05 in every case (99% were under 1.01), suggesting that all chains successfully converged (Gelman and Rubin 1992).
We were interested in evaluating two potential trade-off scenarios: isolation of individual host-use traits, and divisions of host-use traits into clusters (characterized by positive interactions within clusters and negative interactions between them). To characterize isolation of individual host-use traits, we calculated the mean correlation of the ten pairwise comparisons involving each host-use trait (separately for phylogenetic and residual correlations). To characterize associations among groups of host-use traits, we found the network structure that maximized the structure score for the entire network of host-use traits, defined as the sum of all pairwise correlations between host-use traits in the same cluster minus the sum of all pairwise correlations between host-use traits in different clusters (See Supplementary Materials for details). We tested the statistical significance of the resulting values by producing a series of 100 null datasets that simulated independent Brownian motion of a continuous character for performance on each host order along the insect phylogenies, plus additional normally distributed residual variation. We converted the resulting continuous host performance values to a binary host presence/absence character by assuming that only the insect species with the highest performance values for each host taxon were present on that host, with the threshold set by matching the number of species using that host in the empirical data. We then calculated all pairwise correlations between use of the focal hosts, mean correlations per host-use trait, and whole-network structure as we did for the empirical data. Empirical individual host-use trait mean correlations were considered statistically significant when their absolute values exceeded the maximum absolute values of any individual mean in 95% of null datasets. Empirical network structures was considered statistically significant when their scores exceeded the network structure scores of 95% the null datasets.
Acknowledgements
We thank D. Moen, L. Doubleday, and S. Noda for comments that improved the manuscript. This work was supported by the National Science Foundation (EF-1115191 and DEB-1258001)





{kind=link}
{kind=link}
{kind=link}