

ABSTRACT
Novel developmental programs often evolve via the cooption of existing genetic networks. To gain insight into this evolutionary process, we explored an independent cooption event of the TAS3 tasiRNA pathway, which has been repeatedly redeployed over the course of plant evolution, in the moss Physcomitrella patens. We find an ancestral role for this pathway in the spatial regulation of a conserved set of Auxin Response Factors. In moss, this results in stochastic patterning of the filamentous protonemal tissue. Through a combination of computational modeling and experimentation, we demonstrate that tasiRNA regulation confers sensitivity and robustness onto the auxin response. The loss of auxin sensitivity is paralleled by decreased developmental sensitivity to nitrogen, a key environmental signal. We propose that the properties lent to the auxin response gene regulatory network by tasiRNAs, along with their ability to stochastically modulate development in response to environmental cues, have contributed to the tasiRNA-ARF module’s repeated cooption over the course of plant evolution.
INTRODUCTION
The evolution of novel forms is frequently driven by the cooption of existing developmental gene regulatory networks (GRNs) (Erwin and Davidson, 2009). Certain networks appear especially prone to such evolutionary repurposing, leading to the regulation of multiple diverse developmental processes by a single conserved GRN (Carroll et al., 2004a; Plavskin and Timmermans, 2012). The properties that favor the recurring cooption of some GRNs remain largely unknown. One possibility is that frequently coopted GRNs regulate defined cellular processes, allowing the evolutionary redeployment of such ‘differentiation modules’ in a new context (Erwin and Davidson, 2009). On the other hand, it has been postulated that small network motifs may have been repeatedly reutilized by evolution to regulate diverse processes because of their signaling properties (Martinez et al., 2008; Milo et al., 2002).
The mechanism of network cooption has been the subject of extensive investigation, with changes impacting the expression of central GRN components frequently driving the evolution of developmental novelties (Carroll et al., 2004b). Small regulatory RNAs, which play a key role in gene regulation during development in both plants and animals, are one mechanism for driving such evolutionary change (Plavskin and Timmermans, 2012). In addition to regulating the level and spatiotemporal expression pattern of their targets, small RNAs are thought to reduce the inherent noisiness of transcription or, as an outcome of mobility, give rise to sharpened gene expression boundaries (Chitwood et al., 2009; Levine et al., 2007; Schmiedel et al., 2015; Skopelitis et al., 2012). These properties may lend robustness to small RNA-regulated networks, perhaps influencing GRN cooptability. An understanding of how the developmental roles of small RNAs and their targets change over the course of evolution may therefore lead to important insights regarding the properties of GRNs that promote their repeated evolutionary cooption (Plavskin and Timmermans, 2012).
Plants have undergone tremendous diversification since their colonization of land ∼450 million years ago (Kenrick and Crane, 1997). Some notable innovations include the formation of a lignified vasculature, a sporophyte-dominant life cycle, layered meristems, leaves, flowers, fruits, and seed. While these changes in body plan occurred in parallel with repeated losses and gains of developmentally important small RNAs, select small RNA families and their targets have remained conserved since the most recent common ancestor of all land plants (Axtell et al., 2007; Cuperus et al., 2011).The TAS3 trans-acting short interfering RNA (tasiRNA) pathway is in this regard of special interest: while it is conserved throughout land plant evolution, its contributions to development vary extensively even among flowering plants (Plavskin and Timmermans, 2012). TAS3 tasiRNA biogenesis is triggered in response to the miR390-directed cleavage of a set of non-coding TAS3 transcripts, which causes conversion of these precursors into long double-stranded RNAs (dsRNAs) by RNA-DEPENDENT RNA POLYMERASE 6 (RDR6) and SUPPRESSOR OF GENE SILENCING 3 (SGS3). The dsRNA intermediates are subsequently processed by DICER-LIKE 4 (DCL4) into 21-nt tasiRNAs that are phased to yield discrete small RNA species. A subset of these tasiRNAs are biologically active and, like miRNAs, act at the post-transcriptional level to regulate expression of specific gene targets (reviewed in Chapman and Carrington (2007)).
Although related tasiRNA biogenesis pathways initiated by other miRNAs exist (Fei et al., 2013), only the miR390-dependent TAS3 tasiRNA pathway is conserved across land plant evolution (Axtell et al., 2007). Furthermore, only the TAS3 tasiRNAs have demonstrated roles in development. The TAS3-derived tasiARFs, which regulate the expression of AUXIN RESPONSE FACTORS within the ARF3 and ARF4 clade, function in the specification of adaxial-abaxial (top-bottom) leaf polarity in a number of flowering plant species, including Arabidopsis, tomato, rice, and maize (Chitwood et al., 2009; Nagasaki et al., 2007; Nogueira et al., 2007; Yifhar et al., 2012). In addition, these tasiRNAs play a key role in monocot shoot meristem maintenance (Dotto et al., 2014; Nagasaki et al., 2007), contribute to heteroblasty and lateral root outgrowth in Arabidopsis (Hunter et al., 2003; 2006; Marin et al., 2010; Yoon et al., 2010), and regulate leaf complexity and lobing in Lotus and Medicago, respectively (Yan et al., 2010; Zhou et al., 2013). Thus, even within the flowering plant lineage, the TAS3 tasiRNA pathway has been coopted for the regulation of diverse developmental processes.
Moreover, the organs whose development is regulated by TAS3 tasiRNAs in flowering plants - leaves, roots, and layered shoot meristems - evolved long after the pathway itself did (Axtell and Bartel, 2005; Kenrick and Crane, 1997). The genome of the moss Physcomitrella patens, which last shared a common ancestor with flowering plants ∼450 million years ago, includes six TAS3 loci (Arif et al., 2012; Axtell et al., 2007). As in flowering plants, a subset of TAS3-derived tasiRNAs in moss target transcripts of ARF transcription factors (Axtell et al., 2007; Talmor-Neiman et al., 2006). This suggests an additional, yet unknown role of TAS3-derived tasiRNAs in ancient land plant development. Studying the role of these small RNAs outside the flowering plant lineage thus presents a unique opportunity to shed light on potential ancestral functions of this pathway, and to explore the properties that have led to this pathway’s repeated cooption over the course of plant evolution.
To this end, we generated mutants in the moss Physcomitrella patens that lack SGS3 activity and are impaired in tasiRNA biogenesis. The Ppsgs3 mutants exhibit defects in gametophore formation, protonemal branching, and differentiation of specialized caulonemal filaments resulting from the upregulation of a conserved set of repressor ARF genes at the edge of the developing protonema. We show that tasiRNAs act to generate differential levels of ARF expression at the protonemal edge, leading to stochastic patterning of protonemal cell fate determination. Through a combination of computational modeling and experimentation, we further demonstrate that tasiRNA regulation confers sensitivity and robustness onto the auxin response. Finally, we find that Physcomitrella plants defective in tasiRNA biogenesis display decreased developmental sensitivity to a key environmental signal, in line with this pathway’s role in tuning the auxin sensitivity of cells at the plant’s growing edge. We propose that the properties lent to the auxin response gene regulatory network by tasiRNAs, along with their ability to stochastically modulate development in response to environmental cues, have contributed to the repeated cooption of the tasiRNA-ARF module over the course of plant evolution.
RESULTS
Disruption of tasiRNA biogenesis in Physcomitrella impairs protonemal development
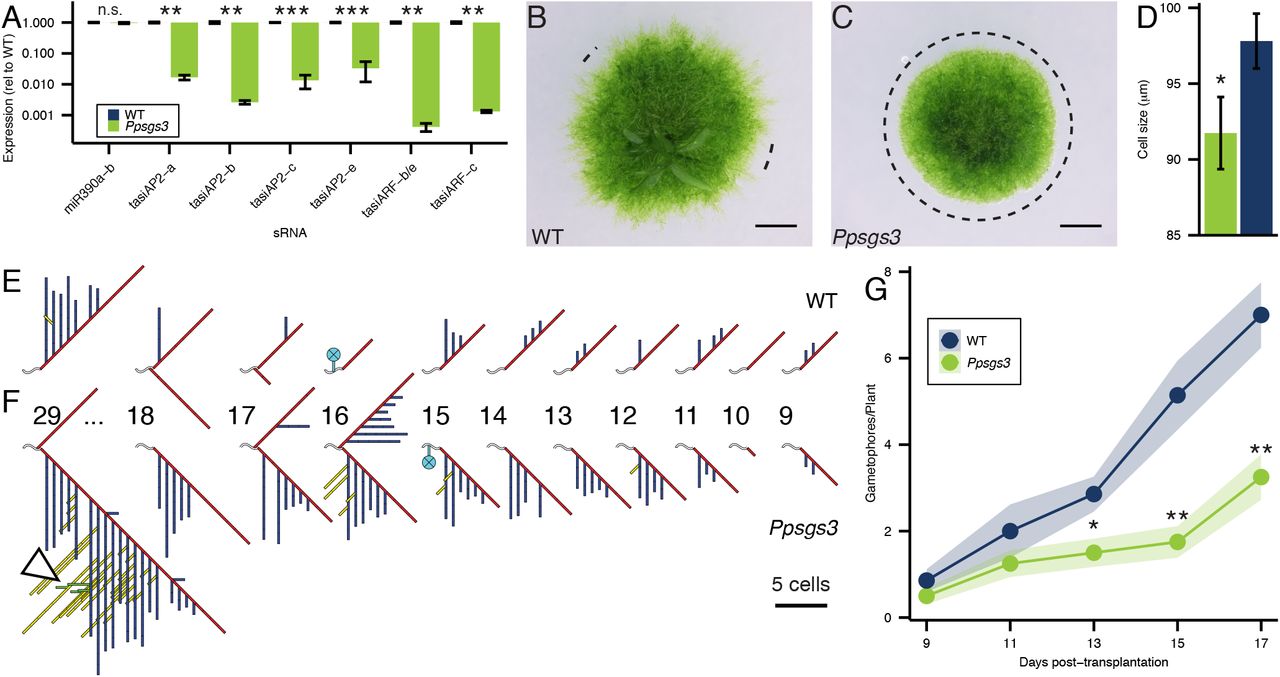
Physcomitrella homologs of RDR6 and DCL4 with a conserved role in tasiRNA biogenesis have been described previously (Cho et al., 2008; Talmor-Neiman et al., 2006). However, the contributions of tasiRNAs to moss development remain unresolved, in part because mutations perturbing these biogenesis components give rise to disparate phenotypes. To gain insight into the developmental role of the TAS3 tasiRNA pathway outside the flowering plant lineage, we used targeted recombination to disrupt the single Physcomitrella homolog of SGS3, PpSGS3 (Figure S1A-E, Table S1). Consistent with a conserved role of PpSGS3 in tasiRNA biogenesis, levels of these small RNAs are drastically reduced in Ppsgs3 mutants (Figure 1A). The Physcomitrella genome includes six TAS3 loci, PpTAS3a-f. These, in contrast to their counterparts in flowering plants, yield two sets of biologically active small RNAs: tasiARF, which target transcripts of four ARF genes, PpARFb1-4, and tasiAP2, which regulate expression of members in the APETALA2 (AP2) transcription factor family (Arif et al., 2012; Axtell et al., 2007; Talmor-Neiman et al., 2006). Levels of both biologically active tasiRNAs are drastically reduced in Ppsgs3 mutants, whereas levels of miR390, which functions upstream of SGS3 in tasiRNA biogenesis, remain unaffected (Figure 1A). In addition, the tasiRNA-guided cleavage of target transcripts is specifically affected in Ppsgs3. Expression of the PpARFb genes is regulated by tasiARF as well as the miRNA miR1219 (Axtell et al., 2007). 5’ RACE analysis revealed that while the miR1219-guided cleavage of PpARFb1 and PpARFb4 transcripts is not noticeably affected, the accumulation of tasiRNA-directed cleavage products for both transcripts is strongly reduced in Ppsgs3 plants (Figure S2A). Together with the striking reduction in tasiRNA levels in Ppsgs3, this indicates that a role of SGS3 in tasiRNA biogenesis is conserved across land plants.

(A) Small RNA levels (mean ± SE, n %3) normalized to wild-type shows that in contrast to miR390, TAS3-derived tasiRNA levels are significantly lower in Ppsgs3 plants than in wild-type. *P < .05; **P < .01. (B-C) Compared to wild-type (B), 15 day-old Ppsgs3 (C) plants form drastically fewer caulonemal filaments. Dotted lines, areas at the protonemal edge lacking caulonema. Scalebar, 1mm. (D) Chloronemal cells in Ppsgs3 filaments are shorter than in wild-type. P < 0.05; n > 20 cells. (E-F) Diagrams of branching patterns along primary filaments from 3 week-old wild-type (E) and Ppsgs3 (F) plants shows a loss of branch determinacy in Ppsgs3. Diagrams show branching patterns at cells 9-18 as well as cell 29 from the tip of the primary filament. Wavy lines, cells of the main filament; numbers, cell position relative to the growing tip; red lines, primary branches; blue lines, secondary branches; yellow lines, tertiary branches; green lines, quarternary branches (white arrowhead); cyan circles, gametophore buds. Branch lengths are proportional to the number of cells in the branch. (G) Gametophore numbers (mean ± SE, n %6) are decreased in Ppsgs3 compared to wild-type. *P < .05; **P < .01.
Loss of PpSGS3 activity results in defined developmental defects. While Ppsgs3 plants show no obvious sporophyte defects and produce viable spores (Figure S2D, E), protonemal development in the mutant is perturbed. Following germination, wild type moss spores produce filamentous, branching protonema comprised initially of chloroplast-rich chloronema. However, as development progresses, or in response to certain environmental cues, dividing chloronemal cells may transition to give rise to the more elongated caulonemal cells. In addition, modified protonemal side branches also give rise to buds, which in turn develop into leafy gametophores (Reski, 1998). While indistinguishable from wild type early in development, the chloronemal networks of 15-day-old Ppsgs3 mutants are smaller and denser than those of wild type (Figure 1B, C). These phenotypes appear to result in part from a decrease in cell size, as chloronemal cells in Ppsgs3 plants are reduced in length by approximately 6% compared to wild type (Figure 1D). In addition, the chloronemal filaments in Ppsgs3 show a more complex branching pattern (Figure 1E, F; S2B, C). Wild type protonemal branching largely follows a stereotyped pattern, in which a ‘main filament’ extending from the center of the plant to its periphery produces one or occasionally two branches at nearly every cell (Figure 1E). In addition, a subset of cells along these primary branches form secondary branches. However, tertiary branches are rarely observed, demonstrating that protonemal branching is effectively determinate. In contrast, Ppsgs3 protonemal filaments consistently show higher order branching patterns compared to wild type protonema of a similar age (Figure 1F; S2C). Both the number of secondary branches and their length, measured as the number of cells per branch, are variably increased in Ppsgs3. Likewise, the number and length of tertiary branches is increased, and quaternary branches regularly occur near the center of 20 day-old Ppsgs3 plants. These data suggest that loss of SGS3 activity and tasiRNA biogenesis causes a decrease in both chloronemal cell size and branch determinacy.
The most striking phenotype of Ppsgs3 mutants, however, concerns the formation of long caulonemal filaments. Under the growth conditions used, long caulonemal filaments visibly extend from the chloronemal edge of wild type plants after approximately two weeks of development, causing older plants to take on a fuzzy appearance (Figure 1B). Although cells with diagonal cross-walls reminiscent of caulonema are occasionally observed in the protonemal network of Ppsgs3 plants, these mutants either completely lack long caulonemal filaments, or rarely form small isolated patches of such filaments along the otherwise smooth protonemal edge (Figure 1B, C). Even after two months of growth, Ppsgs3 plants rarely form long caulonemal filaments, indicating a suppression of caulonemal filament formation rather than a delay in the chloronema-to-caulonema transition upon loss of SGS3 function.
In addition to these protonemal phenotypes, Ppsgs3 plants form significantly fewer leafy gametophores than wild type (Figure 1G), although gametophore morphology itself is normal (Figure S2F-I). This defect is not fully explained by the lack of long caulonemal filaments in Ppsgs3 mutants, as a significant decrease in gametophore number is detected prior to the normal appearance of these filaments at ∼15 days of growth. Indeed, gametophores in Physcomitrella also develop from chloronema (Menand et al., 2007a). Their reduced numbers indicate a role for PpSGS3 in gametophore formation as well as protonemal development. While a reduction in caulonema formation has also been reported for Pprdr6 mutants, the observed decrease in gametophore number in Ppsgs3 is at odds with previous findings showing that Pprdr6 plants develop more gametophores than wild type (Cho et al., 2008; Talmor-Neiman et al., 2006). Potential roles of PpRDR6 outside of tasiRNA biogenesis may account for this difference. PpRDR6 shares a role with PpDCL3 in the production of 22-24 nt siRNAs, and similar to Pprdr6, gametophore production in Ppdcl3 is accelerated (Cho et al., 2008). As Ppdcl3 does not affect tasiRNA biogenesis, a loss of 22-24 nt siRNAs, rather than the loss of tasiRNAs, may account for the accelerated gametophore formation in Pprdr6 as in Ppdcl3.
Taken together, our results indicate that a role for SGS3 in tasiRNA biogenesis is conserved between Physcomitrella and flowering plants. PpSGS3 is required for normal gametophyte development, with Ppsgs3 mutants displaying defects in chloronemal cell size and branch determinacy, caulonemal differentiation, and gametophore formation.
tasiRNAs affect gametophyte development by regulating expression of PpARFb genes
In flowering plants, tasiRNAs exert their effect on development by regulating expression of ARF3 and ARF4 transcription factors (Chitwood et al., 2009; Dotto et al., 2014; Hunter et al., 2006; Song et al., 2012; Yifhar et al., 2012; Zhou et al., 2013). In Physcomitrella, TAS3-derived tasiRNAs similarly target ARF transcripts, and in addition have a unique set of targets in the AP2 transcription factor family (Axtell et al., 2007; Talmor-Neiman et al., 2006). Moreover, an apparent species-specific TAS pathway exists in moss. The Physcomitrella genome includes TAS loci whose transcripts are processed in a miR156-and miR529-dependent manner, referred to as TAS6, which yield tasiRNAs targeting a ZF-domain transcription factor (Arif et al., 2012; Cho et al., 2012). To elucidate which tasiRNA targets are responsible for the developmental defects observed in Ppsgs3 mutants, we compared the expression levels of known tasiRNA targets between wild type and Ppsgs3 in 15-day-old plants, when all aspects of the Ppsgs3 phenotype are apparent. Of the four tasiARF-regulated ARF genes in Physcomitrella, only PpARFb1, PpARFb2, and PpARFb4 are expressed during the gametophyte stage of development (Rensing et al., 2005). In Ppsgs3 plants, transcript levels of all three ARF genes are upregulated ∼2-to 3-fold relative to wild type (Figure 2A). In contrast, expression levels of the tasiAP2 and tasiZF targets are not significantly changed between wild type and Ppsgs3 at this developmental stage (Figure 2A). The observed increase in PpARFb1, PpARFb2, and PpARFb4 transcript levels suggests that the Ppsgs3 developmental defects result, at least in part, from a failure to correctly regulate these tasiARF targets.

(A) qRT-PCR values in Ppsgs3 (mean ± SE, n %3) normalized to wild-type show only transcript levels of the tasiARF targets PpARFb1, PpARFb2, and PpARFb4 are significantly increased in Ppsgs3. *P < .05; **P < .01; ***P < .001. (B-E) Relative to 15 day-old PpARFb4-GUS plants (B), which form near-normal levels of caulonemal filaments, PpARFb4-GUS-m* (C) and PpARFb4-GUS-t* (D) plants show a variably reduction in caulonema number, and PpARFb4-GUS-m*t* (E) plants, with both the miR1219-and tasiARF-complementary sites mutated, only occasionally form caulonema. Dotted lines, protonemal regions lacking caulonema. Scalebar = 1mm. (F) Relative to PpARFb4-GUS, gametophore formation in PpARFb4-GUS-m*t* mutants is severely delayed. Mean gametophore numbers ± SE from at least 10 plants is shown. *P < .05; **P < .01; ***P < .001. (G) In PpARFb4-GUS plants, reporter activity is restricted to the first 1-3 cells at tips of a small subset of chloronemal filaments. (H-J) Reporter activity is detected in more filaments and in more cells along a filament in PpARFb4-GUS-m* (H) and in PpARFb4-GUS-t* (I). This defect is even more pronounced in PpARFb4-GUS-m*t* (J) plants. Insets show isolated filaments; asterisks, cells expressing PpARFb4-GUS; red asterisks, tip cells. Scalebar = 0.1mm.
To investigate this hypothesis, we generated plants that allow the estradiol-inducible expression of HA-tagged, miR1219-and tasiARF-resistant (m*t*) versions of PpARFb2 and PpARFb4 (Kubo et al., 2013; Figure S3A, B), which represent the two distinct branches of the PpARFb clade (Plavskin and Timmermans, 2012). Even when grown on low concentrations of estradiol, the phenotype of plants overexpressing PpARFb2-m*t* or PpARFb4-m*t* resembles that of Ppsgs3. Such plants form denser protonema with fewer gametophores and suppress the transition from chloronemal to caulonemal development (Figure S3C-I). Similar phenotypes are not observed in plants expressing a GUS-GFP fusion protein upon estradiol treatment (Figure S3C, D). This indicates that PpARFb2 or PpARFb4 overexpression is sufficient to recapitulate the Ppsgs3 phenotype, which lends support to the hypothesis that loss of tasiARF activity and the correct regulation of its PpARFb targets underlies the developmental defects of Ppsgs3 mutants. In addition, this finding reinforces the notion that a defect in the biogenesis of small RNAs other than the tasiRNAs accounts for the increased gametophore numbers in Pprdr6 (Cho et al., 2008; Talmor-Neiman et al., 2006).
To substantiate the above findings and to explore the interaction between tasiARFs and miR1219 in the regulation of PpARFb expression, we generated a translational fusion of the endogenous PpARFb4 gene to the GUS reporter, and introduced mutations that prevent targeting of PpARFb4-GUS transcripts by miR1219 (m*), tasiARF (t*), or both small RNAs (m*t*) (Figure S4A-E, Table S1). PpARFb4 was chosen because its expression is especially sensitive to changes in tasiARF regulation (Figure 2A; S2A). PpARFb4-GUS plants show a subtle decrease in the number of caulonemal filaments but otherwise look phenotypically normal (Figure 2B). Likewise, plants expressing the PpARFb4-GUS-m* or PpARFb4-GUS-t* variants show a variable decrease in the number of long caulonema; however, neither fully recapitulates the Ppsgs3 phenotype (Figure 2C, D). Thus, even though PpARFb4 transcript levels are increased approximately 3-fold in Ppsgs3, loss of tasiARF-mediated regulation of PpARFb4 alone is insufficient to recapitulate the Ppsgs3 phenotype. This suggests that altered expression of multiple PpARFb targets is needed to condition the developmental defects observed in Ppsgs3. Indeed, the phenotypic similarities between PpARFb2-m*t* and PpARFb4-m*t* overexpressing plants (Figure S3E-H) indicates that proteins within this clade have related activities.
Importantly, PpARFb4-GUS-m*t* plants fail to form long caulonemal filaments (Figure 2E), closely mimicking the phenotype observed in Ppsgs3. Likewise, PpARFb4-GUS-m*t* mutants show a significant and very strong decrease in gametophore number, with the onset of gametophore formation delayed approximately one week relative to plants expressing the small RNA-sensitive PpARFb4-GUS fusion (Figure 2F). These results indicate that tasiARF and miR1219 coordinately regulate PpARFb4 expression during gametophyte development. Moreover, the phenotype of Ppsgs3 mutants, in which expression of three PpARFb genes is increased due to loss of tasiARF activity, can be phenocopied by perturbation of tasiARF and miR1219 regulation of PpARFb4 alone, as well as by the inducible overexpression of a similar mutant version of PpARFb2 (Figure S3E, F). It follows that tasiARF, in concert with miR1219, regulates the chloronema-to-caulonema transition and gametophore formation in Physcomitrella by fine-tuning the combined level of PpARFb expression. When PpARFb levels overall exceed a certain level, not reached in PpARFb4-GUS-t* plants, formation of long caulonemal filaments and gametophores is inhibited.
tasiARF and miR1219 limit PpARFb4 expression at the protonemal edge
Small RNAs have diverse contributions to plant development (reviewed in Benkovics and Timmermans (2014) and Skopelitis et al. (2012)). Small RNAs are thought to refine levels of protein accumulation by dampening the noise in target gene expression, and can limit expression of their developmental targets to defined spatial and/or temporal domains. Considering that the formation of caulonema and leafy gametophores is in part temporally regulated (Bopp, 1980), tasiARF and miR1219 might control the development of these tissues by regulating the temporal window of PpARFb expression. This possibility seems intriguing, as tasiRNAs in Arabidopsis are known to regulate developmental timing by delaying the expression of adult leaf traits (Hunter et al., 2006). To assess the possibility that tasiARF and miR1219 exert their effects by regulating the timed repression of PpARFb activity, we monitored their accumulation over 3 weeks of gametophyte growth. Protonemal tissue grown on cellophane plates is commonly used to quantify transcript abundance in moss (for example, see Arif et al. (2012); Cho et al. (2008); Pires et al. (2013)), however, such growth conditions substantially impact gene expression levels (Figure S5A, B). Therefore, target gene expression was assayed in intact plants grown under the same conditions used for phenotypic analysis. While expression levels for tasiARF, miR1219, and miR390 are temporally regulated and increase up to ∼60 fold during that developmental time window (Figure S5A), transcript levels for PpARFb1, PpARFb2, and PpARFb4 change comparatively little during that time (Figure S5B).
This observation argues against a role for tasiARF and miR1219 solely in the temporal regulation of PpARFb genes, and instead suggests that these small RNAs may act to maintain a spatial domain of PpARFb expression. To examine this possibility, we compared the pattern of PpARFb4-GUS activity in plants expressing the wild type or small RNA-resistant variants of this reporter. When regulated by tasiARF and miR1219, PpARFb4-GUS expression is limited to 1-3 cells nearest the filament tip at the outer edge of the protonema. Expression in these cells is punctate, consistent with PpARFb proteins functioning as nuclear-localized transcription factors (Figure 2G). Interestingly, PpARFb4-GUS expression is not seen in all chloronemal filaments, but instead appears patchy, with a subset of filaments along the circumference of the plant showing reporter activity. Considering that increased PpARFb expression represses caulonemal filament formation, stochasticity in PpARFb4 levels along the protonemal edge may be linked to the sporadic nature of caulonemal filament formation, which initially appears in seemingly random patches at the edge of the protonema (e.g. Figure 2B).
Mutation of either the tasiARF or the miR1219 target site results in increased PpARFb4 expression, with a greater number of chloronemal filaments and a greater number of cells per filament showing reporter activity (Figure 2H, I). Consistent with the stronger phenotype of PpARFb4-GUS-m*t* plants (Figure 2E), reporter activity is expanded even further upon mutation of both small RNA target sites. In these mutants, PpARFb4-GUS expressing cells occur in nearly every chloronemal filament, and expression extends further down the filament than in either the wild type, PpARFb4-GUS-m* or PpARFb4-GUS-t* lines (Figure 2J). In addition, PpARFb4-GUS levels in individual cells appear stronger than in other genotypes. Importantly, the pattern of PpARFb4 expression in all genotypes does not change substantially from 8 to 15 or 22 days of growth, supporting the hypothesis that tasiARF and miR1219 do not establish a temporal pattern of PpARFb expression in protonema. Instead, the data shows that tasiARF and miR1219 act coordinately along the chloronemal filament to limit the spatial expression domain of PpARFb4 to the outer edge of the growing protonema. Considering that PpARFb1 and PpARFb2 expression also increases in Ppsgs3, it seems likely that tasiARF and miR1219 regulate these targets similarly.
Together, these data provide a basis for the caulonemal defect of Ppsgs3 mutants, and suggest a mechanism by which tasiARF, in concert with miR1219, regulates the chloronemal-to-caulonemal transition. Caulonema are thought to differentiate from chloronemal tip cells (Bopp, 1980). tasiARF and miR1219 regulate this process by limiting expression of PpARFb transcription factors, which act as repressors of caulonemal differentiation, to the outer edge of growing protonema. The combined activity of these small RNAs appears finely balanced such that it generates a randomly variegated pattern of PpARFb expression at the protonemal edge. In a subset of chloronemal filaments, PpARFb activity is depleted even in the tip cell, allowing their differentiation into caulonema. Upon loss of small RNA regulation, whether by disruption of tasiRNA biogenesis or small RNA complementarity within PpARFb transcripts, expression of these ARF proteins persists in a broader domain that encompasses a larger number of chloronemal tip cells, preventing their differentiation into caulonemal filaments. Thus, the contribution of PpSGS3 to Physcomitrella gametophyte development lies primarily in the production of tasiARF and the downstream regulation of PpARFb genes. Interestingly, our data suggests that an absence of PpARFb expression is not sufficient for caulonema differentiation, as a variegated pattern of PpARFb4-GUS activity at the protonemal edge is also observed in young plants not initiating caulonemal filaments. Rather, it appears that tip cells lacking PpARFb expression are competent to differentiate into caulonema, but only do so upon receipt of a distinct signal.
tasiRNAs target repressor ARF transcripts to modulate the auxin response
ARF transcription factors, including the tasiARF targets, form part of a highly conserved GRN that regulates the response to the phytohormone auxin. This network integrates auxin perception into development by controlling the transcription of auxin-responsive genes (ARGs). ARF proteins, which have been classified as either “activator” or “repressor” based on their effect on ARG expression, bind the promoters of such genes in an auxin-independent manner. Transcription factor activity of activator ARFs is, however, blocked in the absence of auxin through dimerization with Aux/IAA proteins. The latter are degraded in response to auxin, resulting in the auxin-dependent derepression of ARG expression (Finet and Jaillais, 2012). Interestingly, the Aux/IAA genes are themselves transcribed in response to auxin signaling, forming a negative feedback loop in the auxin response GRN that is conserved between moss and flowering plants (Middleton et al., 2012; Prigge et al., 2010). The repressor ARFs add additional complexity to this network by competing with activator ARFs for binding to ARG promoters; in flowering plants, this competition between activator and repressor ARFs is thought to allow for the differential regulation of the transcriptional auxin response in space and time (Vernoux et al., 2011).
As in flowering plants, auxin regulates a diverse set of developmental processes in moss, including those impacted by the tasiRNA pathway (Aoyama et al., 2012; Bopp, 1980; Prigge et al., 2010). Indeed, both Ppsgs3 and lines misexpressing the PpARFb targets mimic phenotypes resulting from the treatment of Physcomitrella with the ‘anti-auxin’ compound p-Chlorophenoxyisobutyric acid (PCIB) (Figure S6A, B). The phenotypes of these mutants are also similar to those of classical auxin-insensitive mutants in moss, which form densely packed chloronema, lack caulonemal filaments, and develop fewer or no gametophores (Prigge et al., 2010). These similarities suggest that the PpARFb proteins may act as repressors of the auxin response. This hypothesis is supported by phylogenetic analysis, which places the tasiRNA-targeted ARFs in Physcomitrella sister to the ‘B’ group of ARF genes in flowering plants that includes known repressor ARFs, such as the tasiARF target ARF3 (Plavskin and Timmermans, 2012). Indeed, consistent with a role of the Physcomitrella tasiRNA targets in repressing the auxin response, transcript levels of the early auxin response genes PpIAA1a and PpIAA1b (Prigge et al., 2010) are reduced in Ppsgs3 compared to wild type (Figure S6C).
The role of PpARFb proteins as repressors of the auxin response provides additional insight into the stochastic nature of caulonema formation. The tasiARF-and miR1219-generated variation in PpARFb expression at the protonemal edge results in variable responsiveness of chloronemal tip cells to auxin, which is known to promote caulonemal fate. Importantly, the phenotypes of tasiRNA biogenesis mutants in flowering plants also result from changes in the spatiotemporal expression of B-group ARF genes (Chitwood et al., 2009; Dotto et al., 2014; Hunter et al., 2006; Song et al., 2012; Yifhar et al., 2012; Zhou et al., 2013). By regulating the pattern and level of repressor ARF accumulation, the tasiRNA pathway may thus act to modulate the auxin response across spatial domains in the plant; this finding sheds light on a potential ancestral function of this pathway.
Auxin regulation of PpARFb genes creates a second negative feedback loop in the auxin response network
Considering the conserved role of the tasiRNA pathway in regulating canonical repressors of the auxin response, an understanding of how tasiRNAs and their targets affect the signaling properties of the ancient auxin response network at the cellular level may elucidate potential reasons for its repeated evolutionary cooption. A correct model of the architecture of the auxin response GRN is, in this regard, key. An important conserved feature of this network is the negative feedback between Aux/IAA genes and the auxin response (Prigge et al., 2010). In Arabidopsis, expression of a subset of repressor ARF genes is also subject to feedback from the auxin response (Marin et al., 2010; Paponov et al., 2008; Vanneste et al., 2005; Yoon et al., 2010). As this may impact a GRN’s signaling properties, we determined whether auxin signaling affects expression of the PpARFb tasiRNA targets in Physcomitrella by analyzing PpARFb1, 2 and 4 transcript levels in plants grown on media containing 0.1 μM NAA. Transcript levels for all three genes are increased ∼2.5-fold in auxin-grown plants (Figure 3A). Consistent with this finding, PpARFb4-GUS expression is expanded in plants grown on auxin-supplemented media, with more protonemal filaments and more cells per filament showing reporter activity (Figure S7C, E). Increased expression in response to auxin is also observed for the tasiRNA-resistant form of PpARFb4-GUS (Figure S7D-F), suggesting that auxin promotes PpARFb expression at the transcriptional level, rather than exclusively via repression of tasiARF species. Indeed, the small RNAs regulating PpARFb expression show a complex response to auxin treatment. Although some tasiARF species are upregulated in plants grown on media supplemented with auxin, other tasiARF species, as well as miR1219, are repressed (Figure S4G). These data establish the existence of a negative feedback loop between auxin and repressor PpARFb levels that, in addition to the highly conserved negative feedback between auxin signaling and Aux/IAA genes, modulates the Physcomitrella auxin response GRN. Interestingly, this network configuration appears to be partially conserved, as the Arabidopsis tasiARF target ARF4 is also upregulated in response to auxin (Marin et al., 2010; Paponov et al., 2008; Vanneste et al., 2005; Yoon et al., 2010).

(A) Transcript levels (mean ± SE, n %3) normalized to expression levels in plants grown on media without added NAA shows expression of PpARFb1, PpARFb2, and PpARFb4 is upregulated 2-3-fold in 15 day-old plants grown on 0.1μM NAA. *P < .05; **P < .01; ***P <.001. (B) Schematic of the computational model of the Physcomitrella auxin response network; for details on the parameters and ODEs governing the behavior of the model, see Table SM1 and Equations S1-S9, respectively. i. The model incorporates activating (ARF+) and repressing (ARF-) ARFs, as well as Aux/IAAs, which are subject to auxin-dependent degradation. ii. Aux/IAA and ARF-transcripts are produced through the activity of a promoter containing an Auxin-Responsive Element (ARE). Transcription only occurs when the promoter is bound by an ARF+ monomer or ARF+ dimer, such that the gene is repressed in all other promoter-bound states, as well as the unbound state. The activity of the promoter governs the formation of Aux/IAA and ARF-transcript. ARF-transcript degradation rate or translation rate can be increased or decreased, respectively, to simulate tasiRNA activity. (C) Simulation of ARG expression in response to a prolonged, steady auxin signaling input under both ‘wild-type’ and ‘Ppsgs3’ conditions plotted relative to the pre-signal state, shows ‘Ppsgs3’ has a lower steady-state response to an auxin input. Note the different scales used to plot auxin input and ARG expression. (D) Simulation of ARG expression in response to a noisy auxin signaling input, plotted relative to the steady-state levels of ARG expression in both ‘wild-type’ and ‘Ppsgs3’ conditions, or relative to mean auxin level in the case of the auxin trace. The same noisy auxin signal causes smaller deviations from steady-state levels in ‘Ppsgs3’ than in ‘wild-type’, suggesting the former may be more robust to fluctuations in auxin signaling. (E) Increasing the degradation rate of repressive ARF transcripts to reflect small RNA regulation results in increased sensitivity, but also increased noise amplification, across a large range of γA-transcript values, followed by a small dip in noise amplification. Noise amplification shown with standard error; γA-transcript values for ‘wild-type’ and ‘Ppsgs3’ used in (C) and (D) are marked. (F) Loss of tasiARF regulation increases noise in ARG expression levels. Expression noise increases >2-fold in Ppsgs3 for all ARGs tested except PpRSL1. Across all six ARGs tested, CV is significantly higher in Ppsgs3 than in wild-type (P < 0.05).
Repressive feedback from the tasiARF-targeted ARFs onto the auxin response may impart important properties to the auxin response GRN in Physcomitrella. Negative autoregulatory circuits are central in allowing cells to buffer intrinsic noise, for example resulting from bursts of transcription of circuit components (Alon, 2007; Raser and O’Shea, 2005). However, data from flowering plants has shown that extrinsic fluctuations in auxin signaling input levels may be a significant source of noise in the auxin response GRN (Vernoux et al., 2011). To explore how the feedback loop between auxin signaling and tasiRNA-regulated ARFs affects the propagation of extrinsic auxin noise through the auxin response GRN in Physcomitrella, we adapted a computational model of the auxin response based on ordinary differential equations (ODEs) generated by Vernoux et al. (2011), and modified it to reflect the architecture of the moss auxin response GRN (Figure 3B; Modeling Supplement). Two important measures of GRN function, susceptibility and noise amplification, were considered with and without transcriptional regulation of repressor ARFs by auxin signaling.
Susceptibility (elsewhere also described as sensitivity or gain) measures the change in a system’s output relative to a small change in input (Bintu et al., 2005; Hornung and Barkai, 2008). In this case, susceptibility reports the degree to which ARG expression changes in response to a sustained 1% change in auxin signal, and was assayed via numerical solutions of steady-state ARG transcript values.

Noise amplification, on the other hand, measures the ratio between output noise and input noise (Hornung and Barkai, 2008). Here, it is indicative of the degree to which short fluctuations in auxin signaling levels (extrinsic auxin noise) are translated to fluctuations in ARG expression levels.

Noise amplification was assayed in computer simulations of the auxin response GRN. For a detailed discussion of the modeling approach and the parameters used, see the Modeling Supplement.
Feedback regulation from repressor ARFs onto the auxin response results in decreased susceptibility (Figure SM5A), confirming predictions made in simpler genetic networks by both theoretical work (Paulsson, 2004) and simulations (Hornung and Barkai, 2008). Unlike in the case of susceptibility, there is no clear theoretical prediction regarding the effect of negative feedback on extrinsic noise amplification (Paulsson, 2004). Our model predicts that negative feedback from the PpARFb genes on the auxin response minimally affects extrinsic noise amplification. The small decrease in extrinsic auxin noise amplification that is observed is not significant (Figure SM5A). Both findings are robust across a wide range of starting auxin input levels (Figure S5C), and suggest that one advantage of this network architecture may be that it can buffer intrinsic noise without imparting strong amplification of extrinsic noise. Any benefits repressive feedback regulation in the auxin response GRN has in terms of promoting robustness against intrinsic fluctuations in gene expression are, however, coupled with a loss of susceptibility.
tasiARF regulation promotes a robust auxin response
Small RNA regulation is likewise predicted to lend robustness to the output of a GRN. Indeed, a recent study showed that small RNA regulation in mammalian cells suppresses intrinsic fluctuations in target gene expression (Schmiedel et al., 2015). However, the role of small RNA regulation in modulating extrinsic noise amplification is not known. To visualize how tasiRNA regulation affects the cellular response to auxin, we simulated the output of the above auxin response network upon changes in auxin signal levels (see Modeling Supplement) at two repressor ARF transcript degradation rate (γA-transcript) values. The higher value, representing wild type, is based on the degradation rate of tasiRNA-targeted ARF transcripts in wild type Arabidopsis (Narsai et al., 2007). A 3.5-fold lower γA-transcript value, which results in a change in PpARFb level in line with that observed in Ppsgs3, is used to represent a lack of tasiARF regulation. Increasing γA-transcript resulted in a higher response output to a sustained step-increase in auxin signaling level (Figure 3C), reflecting increased susceptibility. In simulations of a noisy signal, increased repressor ARF transcript degradation rates resulted in larger fluctuations in auxin-regulated gene expression (Figure 3D). This hints at an output of the auxin response that is more susceptible and less robust to extrinsic auxin fluctuations in the presence of tasiRNA regulation of PpARFb repressors.
However, while γA-transcript is higher in wild type than in Ppsgs3, precise ARF transcript degradation rates are not known in Physcomitrella. We thus modeled the effect of a spectrum of γA-transcript values on susceptibility and noise amplification. For a wide range of repressor ARF transcript degradation rates tested, increasing γA-transcript results in increased susceptibility to a small, sustained auxin signal that is correlated with an increase in extrinsic noise amplification in the auxin response GRN (Figure 3E). Substantiating these findings, modulation of ARF translation rates (πA-), which may also be affected by tasiRNA regulation, has the same linked effect on susceptibility and noise amplification levels (Figure SM4). In addition, these outcomes are qualitatively robust to changes in all network parameter values (see Modeling Supplement). This generalizes the observation that tasiRNA regulation imparts susceptibility on the auxin response that is coupled with decreased robustness to auxin input noise. However, beyond a γA-transcript value of ∼0.003, the relationship between these network properties change (Figure 3E). The auxin response GRN is too complex for an analysis of parameters contributing to this shift, but it is interesting to note that the best available estimates of network parameter values suggest that the wild type auxin response GRN may function close to this local maximum in susceptibility and extrinsic noise amplification.
These findings further highlight the possible multifaceted contributions of small RNA regulation to GRN properties. While simulations predict that the small RNA-mediated regulation of PpARFb repressor ARFs impairs the plants’ ability to buffer ARG expression levels against fluctuations in auxin signaling input, small RNA regulation is expected to provide robustness against intrinsic noise in repressor ARF levels (Schmiedel et al., 2015). To experimentally assay the effect of tasiARF regulation on noise in the auxin response, we measured the coefficient of variance for transcript levels of six ARGs (Pires et al., 2013; Prigge et al., 2010) across biological replicates of wild type or Ppsgs3. Transcript levels for five of these genes, PpIAA1a, PpIAA1b, PpIAA2, PpRSL5, and PpRSL6, are more variable in Ppsgs3 than in wild type (Figure 3F), and this effect is significant when measured across all six ARGs. This demonstrates that in Physcomitrella, the overall effect of tasiARFs on the auxin response GRN is to increase robustness of auxin-regulated gene expression, and seems to hint at a distinct primary source of noise in the Arabidopsis and Physcomitrella auxin response GRN (Vernoux et al., 2011). As discussed above, autorepression by PpARFb genes may likewise buffer intrinsic fluctuations in the auxin response.
tasiARF regulation promotes phenotypic sensitivity to auxin
While susceptibility, which reports the change in response output resulting from a minute change in signaling input level (Bintu et al., 2005; Hornung and Barkai, 2008), is a useful measure of network signaling properties, larger changes in input levels more accurately reflect auxin signaling in vivo (Petersson et al., 2009). We therefore next modeled the qualitative effect of tasiRNA regulation on steady-state ARG expression across a broad range of increasing auxin signaling levels. As before, calculations were performed at two γA-transcript values, representing wild type (high γA-transcript) and Ppsgs3 (low γA-transcript), respectively. For both values, the output of the simulated GRN to increasing auxin-signaling levels follows a sigmoid curve. However, the ARG transcript level is lower overall in the Ppsgs3-like regime, with differences between the two regimes especially pronounced at high auxin signaling levels (Figure 4A).

(A) The model predicts steady-state ARG transcript levels, normalized to ‘wild type’ levels at baseline auxin signaling input, increase in both ‘wild-type’ and ‘Ppsgs3’ and follow a sigmoid curve with increasing auxin concentrations. Final induction levels of ‘Ppsgs3’ are much lower than those of ‘wild-type’. The x-axis shows the value of auxin signaling input added to the baseline level in the model (see Modeling Supplement). (B-G) Relative expression (mean ± SE, n %3), normalized to levels in wild-type plants grown on media without added NAA, shows PpIAA1a (B), PpIAA1b (C), PpIAA2 (D), PpRSL1 (E), PpRSL5 (F), and PpRSL6 (G) transcript levels in 15 day-old wild-type and Ppsgs3 plants increases with increasing NAA concentration and generally follows a sigmoid form. Expression of all genes increasing strongly above 0.03 – 0.1 μ NAA. *P < .05; **P < .01; ***P < .001. (H-S) 15 day-old wild-type (H-M) and Ppsgs3 (N-S) plants grown on media supplemented with 0, 0.01, 0.03, 0.1, 0.3, or 1.0 μM NAA. In wild-type, caulonemal growth is strongly induced by NAA concentrations as low as 0.01 μM (I). As auxin concentration is increased, more and more caulonema are formed (H-M). On 0.3 μM NAA and higher, chloronema in the primary protonemal network begin to be converted into caulonema as well (L and M), as evidenced by lighter filament color; protonemal density also notably decreases. Ppsgs3 plants grown without auxin form few or no caulonema (N). Unlike wild-type plants, caulonemal differentiation is not induced by 0.01 or 0.03 μM NAA (O-P). However, higher auxin concentrations partially rescue this defect, and Ppsgs3 plants begin to resemble their wild-type counterparts with respect to caulonemal formation and protonemal mat density on 0.3 and 1 μM NAA (R-S). Dotted line denotes the extent of chloronemal-only filaments at the protonemal edge. Scalebar = 1 mm.
To experimentally test the prediction that tasiRNA regulation sensitizes the auxin response, we measured the transcript level profiles for PpIAA1a, PpIAA1b, PpIAA2, PpRSL1, PpRSL5, and PpRSL6 in wild type and Ppsgs3 plants grown on media supplemented with between 0 - 1μM NAA (Figure 4B-G). Very low doses of exogenous auxin not commonly assayed were included in this range to observe the effect of subtle changes in auxin levels on ARG expression. As expected, expression of all six genes in wild type increases upon treatment with NAA. Consistent with computational predictions, the PpIAA gene expression profiles follow a sigmoidal curve, with the steepest increase in auxin-induced expression occurring between 0.1 and 0.3μ NAA. The change in expression for the PpRSL genes appears more gradual, perhaps reflecting the complex transcriptional interaction between RSL family members (Pires et al., 2013).
Importantly, Ppsgs3 plants display reduced sensitivity to auxin, as predicted by the model. Despite increased variability in transcript levels in Ppsgs3, significant differences between wild type and Ppsgs3 can be observed even at low auxin levels. Transcript levels for all six ARGs are significantly upregulated in response to treatment with auxin concentrations greater than 0.1 μ NAA, but remain lower than those seen in wild type. These results support the prediction made by our computational model that regulation of the PpARFb repressor ARFs by tasiARF results in a dramatically increased sensitivity to changes in steady-state auxin levels that promotes the auxin-dependent induction of ARGs.
The decreased sensitivity of Ppsgs3 plants to auxin at the molecular level is reflected at the phenotypic level. As described above, 15 day-old wild type plants grown without exogenous auxin produce multiple caulonemal filaments around the circumference. The number of these filaments increases visibly in wild type plants grown on 0.01-0.1 μM NAA, with caulonema emerging along most of the protonemal circumference upon treatment with 0.1 μM NAA (Figure 4I-K). On auxin concentrations beyond that, the protonemal network is increasingly comprised of caulonema, resulting in a sparser and lighter appearance (Figure 4L, M) In contrast, Ppsgs3 plants grown on auxin concentrations up to 0.1 μM NAA show a minimal induction of caulonemal differentiation, and continue to form dense chloronemal networks with few or no caulonemal filaments (Figure 4O, P). Treatment with levels of NAA over 0.1 μM, however, can override this defect, such that caulonemal filaments differentiate along the entire protonemal circumference (Figure 4R, S). These findings indicate that Ppsgs3 plants are not generally impaired in their ability to form caulonemal filaments or respond to exogenous auxin. Rather, Ppsgs3 mutants display a reduced auxin sensitivity such that a smaller proportion of chloronemal cells differentiates into caulonema in response to a given auxin signal.
These phenotypic and gene expression data parallel the computational predictions of single-cell auxin responses and demonstrate that tasiRNAs play a key role in sensitizing the auxin response in Physcomitrella via their regulation of PpARFb genes. By spatially regulating PpARFb expression, tasiARFs, in concert with miR1219, create a zone of variable auxin sensitivity at the protonemal edge, allowing the ratio of caulonemal to chloronemal cells to be tuned in response to increasing auxin levels.
Ppsgs3 plants have reduced sensitivity to environmental cues that guide development
Quantitative regulation of development, such as that observed above in the Physcomitrella auxin response, is especially important in plants, which must constantly integrate a wide range of environmental signals into their developmental programs. Caulonemal differentiation appears to be especially labile to environmental regulation, and is modulated by light quality, as well as by the levels of nutrients such as calcium, phosphorus, and nitrogen (Reski, 1998). We hypothesized that, as quantitative regulators of caulonemal differentiation, tasiARFs may play a key role in the interaction between moss and its environment. To test this possibility, we analyzed the effect that reduced substratum nitrogen has on wild type and Ppsgs3 protonemal morphology. For wild type, decreasing nitrogen levels in the growth medium from the typical 5 mM di-Ammonium Tartrate (dAT) promotes the chloronemal-to-caulonemal transition, such that the proportion of caulonemal filaments on plants grown on media containing 1, 0.2, 0.05, or 0 mM dAT increases substantially (Figure 5A-E). Although the number of caulonemal filaments on Ppsgs3 plants likewise increases with lower nitrogen levels in the media, Ppsgs3 plants grown on 0.2 mM dAT and especially 1 mM dAT develop considerably fewer caulonema than wild type (Figure 5F-J), demonstrating diminished sensitivity to changes in substratum nitrogen levels. These results demonstrate that tasiRNAs are essential for allowing a sensitive developmental response to environmental cues that moss is likely to encounter in nature.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A-J) wild-type (A-E) and Ppsgs3 (F-J) plants grown on BCD media supplemented with 5, 1, 0.2, 0.05, or 0 mM diAmmonium Tartrate (dAT) as an additional nitrogen source. Wild-type plants respond to decreased substrate nitrogen content by increasing caulonemal formation. Although Ppsgs3 plants respond similarly to wild-type on media with very low levels of dAT or without any dAT (D-E, I-J), the caulonemal response is either decreased or completely absent in Ppsgs3 plants grown on 1 and 0.2 mM dAT (G, H). Dotted line denotes the extent of chloronemal-only filaments at the protonemal edge. Scalebar: 1mm. Concentrations labeled in terms of Ammonium ion concentrations. (K-N) Following ∼2 months growth on soil, wild-type plants (K, L) often have ‘satellite’ gametophores away from the main plant (approximate edge of main plant highlighted with white dotted line in K and M, gametophores outside of dotted line are satellites; in schematics in L and N, circle represents position of main plant, with asterisks denoting positions of satellite gametophores). Ppsgs3 plants (M, N) form far fewer satellite gametophores and have a more compact plant size. Scalebar: 1cm.
Physcomitrella’s natural habitat is, however, vastly different from the agar plates on which moss is grown in the lab, and includes agricultural fields and the moist soil at the edges of bodies of freshwater (Cuming, 2011). To test how tasiRNA regulation impacts moss development in conditions more reflective of Physcomitrella’s natural environment, we compared the growth of wild type and Ppsgs3 in moist soil. While it was not possible to directly observe protonemal development on soil, two months after transplantation to soil, wild type Physcomitrella have a central cluster of gametophores, ∼2 cm wide, surrounded by many satellite gametophores (Figure 5K, L). By contrast, Ppsgs3 plants have a smaller central cluster and few, if any, satellite gametophores (Figure 5M, N). This result suggests that the decrease in caulonemal differentiation caused by the loss of tasiRNA biogenesis severely impacts the ability of the plant to colonize its substratum, highlighting the importance of tasiRNAs in moss’ interactions with its environment.
Our work shows that by spatially restricting a conserved set of Auxin Response Factor (ARF) targets to a subset of cells at the protonemal edge, tasiRNAs modulate moss’ response to the ancient phytohormone auxin. Modeling the effect of tasiARF-mediated ARF regulation predicts that these small RNAs alter the signaling properties of the auxin response gene regulatory network (GRN), sensitizing cells to auxin while also causing increased amplification of fluctuation in auxin signaling inputs. However, loss of tasiRNA biogenesis results in decreased robustness of auxin-regulated gene expression, likely reflecting the ability of small RNAs to buffer transcriptional noise in their targets. Loss of tasiRNA biogenesis also results in decreased auxin sensitivity, as well as decreased sensitivity to changing substratum nitrogen levels.
DISCUSSION
An ancestral role for TAS3 tasiRNAs in modulating auxin-regulated processes via ARFb repressor proteins
Instances of repeated cooption of specific genetic networks to direct new developmental processes are common across evolution (Carroll et al., 2004a; Plavskin and Timmermans, 2012). This study considers network properties that promote recurrent cooption by investigating one often-repurposed genetic pathway, the TAS3 tasiRNA pathway. Although this small RNA pathway likely originated in the common ancestor of all land plants (Axtell et al., 2007), its known roles in flowering plants are in a diverse set of recently evolved processes, including flower, root, and leaf development (see Plavskin and Timmermans, 2012). Our results demonstrate that in the moss Physcomitrella patens, the TAS3 tasiRNA pathway acts in the gametophytic stages of development to modulate gametophore initiation, protonemal branch determinacy, and caulonemal differentiation. The roles of tasiRNAs in protonemal development likely represent an independent cooption event, rather than an ancestral function, as the extensive and complex protonemal network found in Physcomitrella evolved within the moss lineage (Mishler and Churchill, 1984). Nonetheless, the functional targets of the Physcomitrella tasiRNA pathway are conserved. Its effects on gametophyte development are via regulation of B-family repressor ARFs, whereas the functions of novel tasiRNA targets, such as the AP2-and ZF-family transcription factors (Arif et al., 2012; Axtell et al., 2007; Talmor-Neiman et al., 2006), remain unknown and are not immediately apparent from the Ppsgs3 phenotype. Importantly, tasiRNAs in flowering plants likewise affect development via regulation of members of the B-group of ARF genes (Chitwood et al., 2009; Dotto et al., 2014; Hunter et al., 2006; Song et al., 2012; Yifhar et al., 2012; Zhou et al., 2013). These transcription factors in turn modulate the auxin response in space and time by repressing ARG expression (Finet and Jaillais, 2012). Our findings point to an ancestral role of the TAS3 tasiRNA pathway in modulating the auxin response, with the tasiARF-ARFb module repeatedly coopted for diverse developmental roles over the course of plant evolution.
tasiARFs promote sensitivity and robustness of the auxin response
The strong temporal fluctuations in auxin signaling levels observed in flowering plant meristems suggested that the need to buffer this extrinsic noise may be a key factor shaping the organization and evolution of the auxin response GRN (Vernoux et al., 2011). Our findings predict other properties may have driven the tasiARF-ARFb module’s repeated cooption. Our model predicts that negative feedback from repressor ARFb genes onto the auxin response, which is at least in part conserved in flowering plants (Marin et al., 2010; Paponov et al., 2008; Vanneste et al., 2005; Yoon et al., 2010), minimally affects extrinsic noise amplification. Instead, a predicted advantage of this network feature is that it may limit intrinsic noise resulting from the inherent stochasticity of gene expression (Alon, 2007). A key contribution of the TAS3 tasiRNA pathway to the auxin response GRN during Physcomitrella protonemal development may also be to confer robustness onto the auxin response, as loss of tasiRNA regulation was found to increase variation in auxin responsive gene expression. However, this contribution seems to reflect a suppression of intrinsic rather than extrinsic noise in the auxin response GRN, as our model predicts that tasiRNA regulation amplifies the extrinsic noise from fluctuating auxin signaling input levels. On the other hand, a role for small RNAs in repressing intrinsic noise is in line with recent findings in mammalian cells (Schmiedel et al., 2015). Interestingly, Schmiedel et al. (2015) demonstrated that intrinsic noise repression by small RNAs is especially effective when transcripts contain multiple small RNA binding sites, as is the case for the tasiRNA-targeted ARFb genes in moss as well as flowering plants.
The repeated repurposing of the tasiARF-ARFb module over the course of plant evolution suggests that introducing this module into a novel, auxin-regulated context may provide a selective advantage. Both the increased sensitivity and robustness lent to the auxin response by the tasiRNA pathway promote the faithful transfer of information through a signaling network. Evidence that selection may indeed act on such network properties has been identified across the tree of life, and includes examples of selection acting on regulatory mutations that minimize expression noise in yeast (Metzger et al., 2015; Wang and Zhang, 2011), as well as the conservation of shadow enhancers that maintain robust gene expression in animal systems (Frankel et al., 2010; Hong et al., 2008). The network properties lent to the auxin response by the tasiARF-ARFb module uncovered in this work thus provide a compelling explanation for the repeated cooption of this module throughout land plant evolution. Furthermore, these signaling properties may provide an explanation for the prevalence of evolutionary conserved small RNA-target modules in plants as well as animals.
Considering these points, it is interesting to note that a subset of the flowering plant B-group ARF genes have lost tasiRNA regulation (Plavskin and Timmermans, 2012). Our modeling results provide a possible explanation for this diversification. As plants evolved the need to regulate auxin responses in organs where fluctuations in auxin levels are high, such as in meristems (Vernoux et al., 2011), the benefit provided by tasiRNAs in dampening intrinsic expression noise may have been offset by their amplification of extrinsic auxin input noise. Likewise, despite the apparent importance of small RNA regulation in promoting sensitivity and robustness of the auxin response, many auxin-regulated processes in Physcomitrella development, such as in the leafy gametophore and sporophyte (Bennett et al., 2014; Viaene et al., 2014), are unaffected when tasiRNA function is perturbed. An interesting direction for future studies may be to explore whether miR160 and its highly conserved targets in the C-family of repressor ARFs (Axtell and Bartel, 2005; Finet et al., 2012; Plavskin and Timmermans, 2012), act in place of the tasiARF-ARFb module in the regulation of these processes.
tasiARF-mediated PpARFb regulation allows for stochastic protonemal cell fate determination
In addition to the network properties acting at the cellular level, the tasiARF-mediated regulation of PpARFb expression generates stochasticity at the whole plant level. Moss protonema is a heterogenous tissue, with caulonemal filaments specified at seemingly random locations along the edge of the protonemal network over the course of development. Well-known examples of tissues where cell fates are specified using a stochastic choice mechanism to create a randomly mixed population of cell types exist in the metazoan nervous system, and include the diversification of olfactory and visual sensory neurons (Johnston and Desplan, 2010). However, the source of stochasticity in these examples is unclear.
Our observations indicate that tasiARF, in conjunction with miR1219, regulates caulonemal fate specification in Physcomitrella by limiting PpARFb expression to a random subset of chloronemal tip cells. To create variability in PpARFb expression across individual filaments tips, the levels of small RNAs in the protonema must be carefully tuned. In this regard it may be interesting that in Arabidopsis, the cell-to-cell movement of tasiARF from a localized source creates a small RNA gradient across developing leaf primordia (Chitwood et al., 2009). A gradient of tasiARF and miR1219 activity that dissipates towards the filament tips, perhaps generated by small RNA movement from a source at the center of the protonemal network, presents a possible mechanism to generate stochastiocity in PpARFb expression at the protonemal edge. The fact that the number of cells expressing PpARFb varies from filament to filament can also be explained by local subtle variation in the shape of a small RNA activity gradient. If so, patterning by small RNA gradients may represent a novel mechanism of stochastic cell fate specification.
Stochastic cell fate decisions are often important for environmental plasticity, allowing ‘bet-hedging’ of cell fate choices within populations of cells (Johnston and Desplan, 2010). In combination with the network properties small RNA regulation confers on the auxin response GRN, stochasticity in caulonema specification resulting from spatial variation in tasiARF and miR1219 regulation of PpARFb proteins appears important for developmental plasticity. To fine-tune development in response to subtle gradations in environmental conditions, organisms must modulate development in a quantitative, graded manner. However, to create such gradation in a binary system, such as the chloronemal-caulonemal cell fate switch, cells must display variability in their response to the switch-inducing signal (Ferrell and Machleder, 1998). By limiting PpARFb expression to a random subset of chloronemal tip cells, the tasiRNA pathway, along with miR1219, creates variable auxin responsiveness at the edge of the developing protonema. Tip cells with comparatively high small RNA activity and low repressor ARFb levels have increased auxin sensitivity, and are thus more likely to differentiate into caulonema in response to auxin signaling. Our finding that the number of caulonemal filaments increases progressively with increasing auxin levels supports this notion. The decreased response to small changes in substratum nitrogen availability in moss plants defective in tasiRNA biogenesis further underscores the importance of tasiRNAs in maintaining a plastic response to the environment.
With the role of tasiRNAs in sensitizing development to environmental inputs in mind, parallels can be drawn between independently evolved tasiRNA-regulated processes in mosses and flowering plants. Lateral root initiation in Arabidopsis, although likely sharing little with protonemal development in terms of cellular mechanisms, is also auxin-and tasiRNA-regulated, and is carefully tuned by environmental inputs, including substratum nitrogen levels (Gifford et al., 2008). Likewise, the auxin-and tasiRNA-regulated specification of abaxial-adaxial polarity in flowering plant leaves is sensitive to environmental inputs, with the number of adaxial and abaxial cell layers regulated in part by light quality (Kozuka et al., 2011).
Network signaling properties as drivers of GRN cooption
Although the mechanisms by which cooption of genetic networks occurs have been extensively investigated, the reasons for the preferential repeated cooption of select networks are less well understood. One potential benefit of GRN cooption is that it allows the redeployment of a ‘differentiation gene battery’ involved in a specific cellular or developmental process (Erwin and Davidson, 2009). For example, the RSL genes were repeatedly repurposed during the evolution of plant organs that develop via filamentous growth, such as protonema in moss and root hairs in flowering plants (Jang et al., 2011; Menand et al., 2007b; Pires et al., 2013). In contrast, we propose that the repeated cooption of the TAS3 tasiRNA pathway was driven by the properties that this pathway lends to the auxin response, rather than by the redeployment of a specific downstream developmental process. We find that tasiRNAs lend the networks they regulate two key properties: robustness to intrinsic noise in target levels, and sensitivity to environmental signals. Further, small RNAs may provide a mechanism to create stochastic cell fate patterns that are developmentally plastic and increase the spectrum of responses to environmental stimuli. Considering the signaling properties of a genetic network, and not just its developmental output, may thus be critical to understanding the evolution of complex multicellular forms.
MATERIALS AND METHODS
The transformation and propagation of Physcomitrella was performed as described (Cove et al., 2009a, 2009b), with minor adjustments detailed in Supplementary Materials and Methods. For phenotypic or gene expression level measurements, protonema were subcultured on cellophane plates 2-3 times, and small pieces (1-3 mm diameter) transplanted to BCDAT plates containing 0.9 μ FeSO4 at pH 5.8, unless otherwise noted. For chemical treatments, auxin, PCIB, or estradiol was added at the indicated concentrations. Nitrogen level experiments were performed on BCD plates supplemented with the appropriate amount of diammonium tartrate. For soil experiments, plants were grown on Rediearth (Sungro) under a plastic dome for two months at 22°C under a 16/8 light/dark regime. Cell size measurements were performed on chloronemal cells 3-5 cells from filament tips. Branching was analyzed on 3 week-old plants. For, gametophore counts, the number of gametophore buds with at least 1 phyllid were counted. Histochemical staining was performed on 15 day-old plants as described (Chitwood et al., 2009). For all qRT-PCR and 5’ RACE experiments, total RNA was extracted from Physcomitrella using Trizol reagent (Invitrogen) according to the manufacturer’s suggested protocol. Unless otherwise stated, RNA was extracted from whole 15 day-old plants as detailed in Supplementary Materials and Methods. Small RNA qRT-PCR was performed following a protocol modified from Varkonyi-Gasic et al. (2007), and RLM 5’ RACE was performed as described in Axtell et al. (2007). All expression levels were calculated relative to untreated wild-type controls, after normalization to levels of GAPDH (gene expression) or U6 (small RNA expression). Statistical significance was calculated from at least three biological and two technical replicates using student’s t-test between wild-type and mutant samples within a given condition. For detailed experimental procedures, see Supplementary Materials and Methods. For detailed computational modeling procedures, see Supplementary Modeling Information.
AUTHOR CONTRIBUTIONS
YP and MT conceived the study and designed the project. YP and MT planned the experiments with input from P-F.P, AN, MH, and RQ. YP, P-F.P, and AN generated the transgenic moss strains. YP performed the experiments. YP performed the computational modeling with input from GA and MT. YP and MT wrote the manuscript.
ACKNOWLEDGEMENTS
We thank members of the Timmermans, Quatrano, and Hasebe laboratories who have contributed ideas and thoughtful comments to the manuscript, Tim Mulligan for plant care, and Elijah Salome-Diaz, Carol Hu, Cristina Marco, and members of the Hasebe lab for technical assistance. We also thank Teva Vernoux for sharing modeling code. YP was funded by an Al Hershey fellowship from the Cold Spring Harbor Laboratory Watson School of Biological Sciences and a predoctoral training grant 5T32GM065094 from the National Institute of General Medical Sciences. This work was also supported by the East Asia and Pacific Summer Institutes grant OISE-1015679 from the National Science Foundation and the Japan Society for the Promotion of Science. MH is funded by ERATO and KAKENHI from MEXT, Japan. Research on small RNA regulation in the Timmermans lab is supported by grants from the National Science Foundation (IOS-1355018 and MCB-1159098) and an Alexander von Humboldt Professorship from the Deutsche Forschungsgemeinschaft.
REFERENCES




